the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 11 Sep 2025
| 11 Sep 2025
What if extreme droughts occur more frequently? Mechanisms and limits of forest adaptation in pine monocultures and mixed forests in Berlin–Brandenburg, Germany
Jamir Priesner
Boris Sakschewski
Maik Billing
Werner von Bloh
Sebastian Fiedler
Sarah Bereswill
Kirsten Thonicke
Britta Tietjen
Forests in eastern Germany are already experiencing the detrimental effects of droughts, exemplified by the severe conditions of the 2018 drought year. With climate change, such extreme events are expected to become more frequent and severe. Previous work suggests that mixed forests exhibit greater resilience against droughts than monocultures. Our study aims to investigate the impact of increased frequency of extreme droughts, such as those seen in 2018, on biomass, structure, and traits of forests in the eastern German federal states of Berlin and Brandenburg.
Utilizing the flexible-trait dynamic global vegetation model LPJmL-FIT, we simulate the growth and competition of individual trees in both pine monoculture forests and mixed forests. The trees belong to different plant functional types or in the case of pine forests are parametrized as Pinus sylvestris. We create drought scenarios from high-resolution climate input data by re-shuffling the contemporary climate with increased frequencies of the extreme drought year 2018. For each scenario, we simulated vegetation dynamics over 800 simulation years which, allowed us to analyze shorter-term impacts in the first decades of the drought scenarios, as well as the long-term adaptation of the two forest types to those new climate normals. We evaluated the resulting long-term changes in biomass, plant functional traits, and forest structure to examine the new equilibrium state emerging for each scenario.
Our findings revealed nuanced responses to increased drought frequency. In pine monoculture forests, increased drought frequency reduced biomass and increased biomass variance, indicating higher system instability. Conversely, in mixed forests, biomass initially declined in scenarios with increased drought frequency but eventually recovered and even exceeded baseline levels after 100–150 years. We explain recovery and increase in biomass through two forest adaptation mechanisms; first, we saw a shift in the plant community towards broadleaved trees, and second, plant traits shifted towards increased average wood density, decreased average tree height, and increased average tree age. However, for the most extreme scenario with drought occurring each year, the adaptive capacity of the mixed forest was exceeded, and the biomass halved compared to the baseline scenario. In our study, for the first time LPJmL-FIT is used with a resolution as high as 2 × 2 km2, which allows us to observe spatial heterogeneity drought impacts within the Berlin–Brandenburg area. Pine monocultures suffered, especially in the warmer urban areas, and mixed forests in the central–western part of Brandenburg benefitted in the long term.
This study highlights the capacity of natural mixed forests in contrast to pine monocultures to adapt to increasing drought frequency up to a certain limit. The results underscore the importance of considering biodiversity in forest management strategies, especially with regard to more frequent dry periods under climate change.
- Article
(2740 KB) - Full-text XML
- BibTeX
- EndNote




With temperatures rising at about twice the global average rate, Europe is the fastest warming continent (Copernicus, 2023) and is exposed to more intense and frequent climate extremes (Pradhan et al., 2022; Treydte et al., 2024). Within Germany, Brandenburg is one of the driest regions (Dittmann et al., 2024), with a warming of 1.1 °C over the last decades (LfU Brandenburg, 2022) and consequently increasing evaporation and decreasing groundwater recharge (LfU Brandenburg, 2022). Increasing drought severity and frequency have been observed to damage forests in Brandenburg and to increase the severity of wildfires (Land Brandenburg, 2023; LfU Brandenburg, 2021).
Germany was heavily affected by two consecutive drought years in 2018–2019, where the annual precipitation amount was so low that drought effects extended into 2020 (Büntgen et al., 2021). The extremely dry and hot growing season (March–November) in 2018 was record-breaking in both its high temperatures and low precipitation (Zscheischler and Fischer, 2020) and led to a record in burned forest area due to wildfires in Brandenburg (1664 ha of burned area in Brandenburg, Landeskompetenzentrum Forst Eberswalde, 2018). This compound event affected agricultural production, requiring federal states to grant aid payments of EUR 340 million to farmers with at least 30 % yield loss (Reinermann et al., 2019; Buras et al., 2020). During the consecutive drought years the fraction of trees with signs of damage increased from 53 % in 2017 to 92 % in 2022 (Land Brandenburg, 2023). In 2021, 26 % of the forest area covered with pine in Brandenburg (Berlin: 20 %) showed significant damage in their tree crowns, while the area affected in Brandenburg's beech and oak forests amounted to 40 % and 42 %, respectively (Bundesministerium für Ernährung und Landwirtschaft, 2021).
Since medieval times, natural forests have been cleared for agriculture and pastures, with natural or semi-natural forests left in only a few small regions in Europe (Barredo et al., 2021; Bengtsson et al., 2000; Lamentowicz et al., 2020). In Europe, systematic forest management dates back to the 19th century (Niedertscheider et al., 2014). Today, most European forests are highly managed; single-species monocultures are often planted to provide people with timber from highly productive forests. In Brandenburg, pine trees make up 70.1 % of the forest area, followed by oak and beech with only 6.7 % and 3.3 %, respectively (Land Brandenburg, 2023). To act against increasing forest loss from climate extremes and to support forest adaptation to a changing climate, increasing biodiversity has been suggested as one solution. According to the biological insurance hypothesis, ecosystems with high biodiversity can better maintain ecosystem functioning under external pressure. In addition, diverse forests can hold a larger portfolio of plant strategies that can help them to adapt to the new environmental conditions. Monocultural ecosystems, however, lack the required response diversity to maintain ecosystem functioning under changing environmental conditions (Mori et al., 2013; Yachi and Loreau, 1999).
Changing climate conditions can lead to environmental filtering and thus to a shift in the spatial domain where species can occur and be productive. Respective shifts in species' spatial distribution are among the most significant and most widely discussed ways of how forests in the Northern Hemisphere react/adapt to climate change (Astigarraga et al., 2024; Fei et al., 2017; Lenoir and Svenning, 2015; Parmesan and Yohe, 2003; Rabasa et al., 2013; Rubenstein et al., 2020, 2023). Due to the increase in water deficit and temperature, range shifts upward and poleward are expected by ecological theory (Bonebrake et al., 2018; Lenoir and Svenning, 2015). While there are observations of species for which these expected shifts happen (Chen et al., 2011; Lenoir and Svenning, 2015; Parmesan and Yohe, 2003), many species show multiple directions in response to climate change (Fei et al., 2017; Rabasa et al., 2013; Rubenstein et al., 2023; Zhu et al., 2014). In addition to a range shift in species, changes can also occur in stocks that persist at a specific location, particularly with regard to productivity. However, the effects of climate change on European forests remain unclear (Pretzsch et al., 2023). While there is generally a trend toward more productivity in temperate European forests (Charru et al., 2017; Pretzsch et al., 2014, 2023; Zhu et al., 2014), increasing drought events interrupt this trend (Martinez del Castillo et al., 2022; Piovesan et al., 2008; Schmied et al., 2023; Schuldt et al., 2020; Williams et al., 2013). Due to contrasting trends within regions (Galván et al., 2014; Pretzsch et al., 2023) and among species (Martinez del Castillo et al., 2022; Pretzsch et al., 2014, 2020, 2023), understanding of long-term shifts is still lacking.
Building on the knowledge that more diverse forest ecosystems could be more resilient, recent forestry programs in Germany for example aim at increasing deciduous tree cover to adapt forests to future climate change conditions (Land Brandenburg, 2011; Wessely et al., 2024). This is supported by future projections of average decadal changes in forest dynamics and tree species distribution (e.g., Wessely et al., 2024) and how they affect the ecosystem services provided by German forests (Gregor et al., 2022; Gutsch et al., 2018). Recent model applications studied the importance of functional diversity for future forest adaptation (Billing et al., 2022, 2024). However, we still have a limited understanding of the mechanisms and limits of diverse forests to adapt to an increasing frequency of climate extremes as the new climate normals.
In addition to biodiversity and species identity, it is also useful to consider structural and functional plant traits that determine the reactions to environmental factors and their changes but can also influence the functioning of ecosystems (Sterk et al., 2013; Suding et al., 2008). Wood density and specific leaf area might strongly impact species' responses to climate change. Some studies suggest that higher wood density correlates with drier and warmer climate (Nabais et al., 2018; Zhang et al., 2011; Swenson and Enquist, 2007; Nelson et al., 2020; Bouchard et al., 2024). Most notably, in a recent global tree inventory analysis for temperate forests Bouchard et al. (2024) found higher wood density with decreasing rainfall (below values of 1000 mm a−1, which would also apply to climate conditions in Brandenburg). Fei et al. (2017) observed that in the eastern part of the United States of America tree species that shifted to drier areas had higher median wood density. A global meta-analysis of tree mortality in response to drought found that in addition to wood density, specific leaf area (SLA) also explains drought responses, where trees with a lower SLA showed lower mortality responses (Greenwood et al., 2017). Experimental results also show that individuals of different tree species from the Mediterranean area growing under drought stress had a decreased SLA in comparison to individuals of the same species growing in the control (Valladares and Sánchez-Gómez, 2006), showing that this might be a potential adaptation mechanism.
Forests develop and change over decadal timescales, making it difficult to conduct field experiments that assess how climate extremes impact biodiversity–ecosystem functional relationships. Instead, biodiversity-enhanced, process-based vegetation modeling can be applied to projects and explain how climate extremes affect functional trait composition and ecosystem function in diverse forests and compare them against the performance of monoculture forests. However, climate models might underestimate the frequency of hot–dry compound events like the 2018 drought (Zscheischler and Fischer, 2020; van der Wiel et al., 2021) that were much rarer in the past. The realism of the frequency and intensity of such extreme compound events can vary in climate models, so the resulting simulated impacts on vegetation and tree demography might be blurred and miss out on possible abrupt changes. Therefore, we take a simplistic approach of designing climate scenarios with artificially increased drought frequency for the area in Berlin and Brandenburg in Germany. We use these artificial drought scenarios as input data for the flexible-individual trait dynamic global vegetation model LPJmL-FIT (Sakschewski et al., 2015; Thonicke et al., 2020), which simulates functional and structural trait changes in conjunction with ecosystem functions under varying climate and soil conditions. We then analyze how in Brandenburg and Berlin temperate mixed forests and pine monoculture forests (parameterizing Pinus sylvestris trees) perform and adapt to changing frequency of climate extremes. However, forest management is considered in neither forest type, which means that the pine monoculture forest can be regarded as a semi-natural forest. Additionally, we assume that the entire study area is covered by forest to take advantage of the high-resolution climate data and include urban forest areas. In this context, this study aims to answer the following questions:
-
Does a diverse natural forest have a higher resilience against an increased frequency of extreme drought years such as 2018 than a pine monoculture forest?
-
What are the underlying mechanisms that enable forests in Brandenburg to adapt to the increased frequency of extreme droughts? In particular, how do these mechanisms manifest in the shifts in tree community composition and changes in the traits spectrum within individual plant functional types?
-
Is there spatial variability in the response of the two forest types towards droughts across Berlin and Brandenburg?
We first describe how biomass of the pine monoculture vs. temperate mixed forests is changing under the different drought extreme scenarios before we analyze how structural and functional traits explain the underlying mechanisms and how these mechanisms differ between plant functional types (PFTs).
We created artificial climate data sets with increased drought frequencies using high-resolution climate data compiled for the study area Berlin–Brandenburg as the baseline (Bart et al., 2025). Our new drought scenarios contain weather data from 1980–2022, to which we have added the drought year 2018 with varying frequency. We investigated the impact that these scenarios might have on pine monocultures which currently dominate managed forests in the study area and on mixed forests as their natural analog. We applied the flexible individual-trait dynamic global vegetation model LPJmL-FIT to two plant community configurations, (i) a pine monoculture forest and (ii) a mixed forest, and calculated resulting forest development for both a baseline scenario (the original climate data set) and our new drought scenarios (see below). We simulate the study area to be fully covered by vegetation, neglecting land used for settlements and agriculture. Forest management, such as thinning or logging, was not simulated in any of the configurations. We then analyzed changes in vegetation dynamics and in plant characteristics at the centennial timescale to analyze the short- and long-term ability of forests to adapt to an increased frequency of extreme droughts.
2.1 Model description: the flexible-trait DGVM LPJmL-FIT
The dynamic flexible-trait vegetation model LPJmL-FIT (“Lund–Potsdam–Jena managed Land – Flexible Individual Traits”) is a process-based dynamic global vegetation model (DGVM). It simulates the establishment, growth, competition, and mortality of individual trees using a forest gap approach. Tree individuals can differ in their functional traits according to the leaf and stem economics spectrum (Sakschewski et al., 2015; Thonicke et al., 2020). The spatial resolution of model simulations depends on the resolution of the input data. For each grid cell, the model requires soil texture as well as daily climate input data (temperature, precipitation, and radiation) and atmospheric CO2 concentration to calculate soil hydrology and vegetation dynamics. Grid cells are further subdivided into independent forest patches of 10 m by 10 m on which tree individuals compete for water and light. The present study uses the model version as described in Thonicke et al. (2020) and Billing et al. (2024) and has been extensively validated. In addition, we adopted the variable rooting scheme described in Sakschewski et al. (2021) to allow for diverse tree rooting strategies and excluded grass PFTs from our simulations.
Tree individuals are typically categorized into broad PFTs representing main ecological characteristics of natural vegetation at the biome level as in the standard model LPJmL (Schaphoff et al., 2018). However, the model can also be parameterized for specific species. In LPJmL-FIT newly established tree individuals are randomly assigned to PFTs if there is more than one PFT simulated at the same time. Key functional traits, such as specific leaf area (SLA) and wood density (WD), are then randomly and independently sampled out of the PFT- or species-specific ranges and remain constant over a tree's life. Other functional traits (e.g., leaf nitrogen content) are connected to SLA and WD via trade-offs according to the plant economics spectrum. Trees compete for light and water in independent 10 m × 10 m forest patches. Their crown area and leaf area index control their capacity to absorb photosynthetic active radiation. Water uptake depends on root depth and soil moisture availability. The amount of absorbed photosynthetic active radiation, soil water uptake, and other environmental factors such as temperature and atmospheric CO2 concentration determine the gross primary production (GPP) via the process of photosynthesis. Autotrophic respiration is divided into maintenance and growth respiration, both of which are temperature-dependent and linked to the tree's biomass and GPP. Carbon that is lost through autotrophic respiration is subtracted from GPP, resulting in net primary productivity (NPP), which represents the carbon available for new growth. The allocation of NPP to various parts of each individual tree – roots, stems, and leaves – is modeled based on the specific strategies of each PFT to optimize resource use in different environmental conditions (Schaphoff et al., 2018). Over time, performance and competition determine tree survival and growth. Via these processes, climate, soil properties, and competition conditions filter locally best adapted (environmental filtering) and best performing (competitive filtering) tree individuals. That is, LPJmL-FIT can simulate functionally diverse forests but also monocultures that would grow under the sole influence of climate and soil conditions. An illustrative video of forest community assembly is available in Billing et al. (2024), Video 1, and can be found under the following link: https://www.pik-potsdam.de/~billing/video/2023/spinup_LPJmLFIT.mp4 (last access: 25 August 2025). In this animation, each tree is colored according to its SLA or WD value assigned at establishment. SLA and WD are key traits to determine the performance of trees under environmental filtering and competition in our model. Trees with high SLA benefit from high production compared to carbon investment but have higher leaf turnover rates and a lower photosynthetic capacity (Sakschewski et al., 2015). Higher SLA increases the shading of other trees and therefore brings benefits in light competitiveness (Billing et al., 2022). WD also plays an important role in light competition, as lower WD needs less carbon investment and can lead to faster growth. This can increase productivity and therefore decrease mortality in a light-limited environment. On the other hand, trees with higher WD have a lower probability of dying when the annual performance is low because the maximum of growth-efficiency-related mortality is anti-correlated with WD (Sakschewski et al., 2015). For a more in-depth discussion about the trade-offs connected to trait variation see Billing et al. (2024), especially Fig. 5, and supplementary methods in Billing et al. (2022).
There is no explicit drought-mortality function implemented in the model and also no explicit plant hydraulics. Plant-water uptake and photosynthesis are connected via stomatal conductance. If soil water content is very low, transpiration is reduced, affecting photosynthesis and leave phenology, which leads to abscission and limits productivity. The reduced productivity results in low growth efficiency and therefore increases mortality (Thonicke et al., 2020).
2.2 Data and simulation experiments
In our simulations the area of Brandenburg and Berlin was represented by a grid of 7073 cells with ∼ 2 × 2 km resolution. For each grid cell, 80 patches of 10 m × 10 m patch size were simulated, representing the forest of the total grid cell. We ran the model with climate data derived from the Central Europe Refined analysis version 2 (CER v2) (Bart et al., 2025). This data set was generated by dynamical downscaling of ERA5 reanalysis forcing data provided by the European Centre for Medium-Range Weather Forecasts (ECMWF) for the area of Berlin and Brandenburg, utilizing the Weather Research and Forecasting (WRF) model version 4.3.3. The climate data cover the period from 1980 to 2022 with daily temporal resolution and 2 × 2 km spatial resolution. We first created a climate set for a 1000-year model spinup, randomly drawing from the climate input years 1980–2022. To assess the effects of droughts, we manipulated the original climate data set afterwards by artificially adding the drought year 2018 to the data in increasing frequencies. For a slightly wetter scenario (Scenario A), we only used data from 1980–2001 (i.e., frequency of drought year 2018 = 0), and for the baseline scenario (Scenario B, frequency = 0.02), we used the original full data set from 1980–2022 without any manipulation. Five additional scenarios contained the years 2001–2022 plus the drought year 2018 at varying frequencies (Scenario C, 0.05, to Scenario G, 0.68). For the last scenario we only took data from the year 2018 (Scenario H: 1.0). Increasing the frequency of the 2018 drought year also changed the mean climate. To quantify the drought effect, we calculated the mean annual maximum climatic water deficit (MCWD) following Sakschewski et al. (2021). The absolute values and their deviation from the baseline climate as well as the frequency of the year 2018 for each scenario are shown in Table 1. To create the full weather data sets to run the model for different drought scenarios (Scenarios A–H), we randomly draw 800 years from the respective manipulated climate data set (Fig. 1).
Table 1Characteristics of drought scenarios. Frequency of the year 2018 in manipulated data and resulting mean maximum climatic water deficit (MCWD) and mean temperature (T) as well as deviation from baseline (Scenario B) for each of the scenarios.
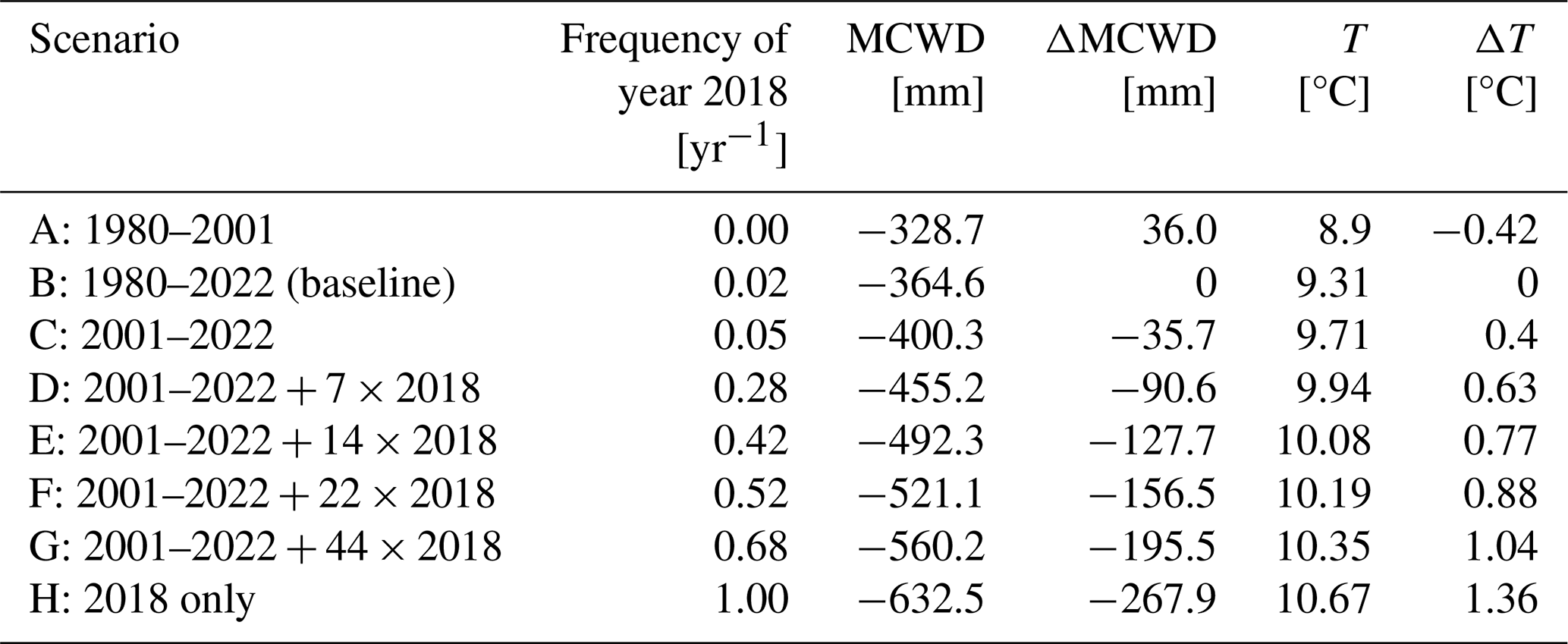
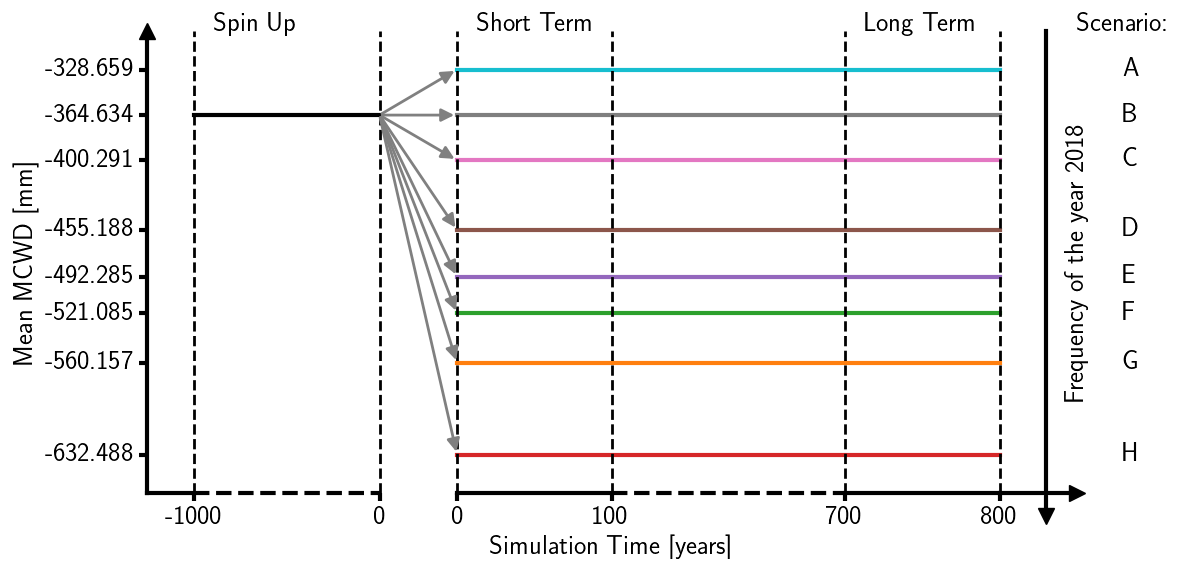
Figure 1Scheme of the simulation protocol. After 1000 years of spinup with shuffled Central Europe Refined analysis version 2 (CER v2) data from 1980–2022, 800 years of Scenarios A–H with decreasing mean maximum water deficit (MCWD) and increasing frequency of the year 2018 follow. The frequency of the year 2018 was increased by adding an increasing number of the year 2018 to the pool from which each year was drawn (see Table 1 for details).
Soil depth data were sourced from Pelletier et al. (2016) and subsequently re-gridded from an original resolution of about 1 km to match the climate data grid resolution. Soil type information was obtained from the Harmonized World Soil Database (HWSD) (FAO and IIASA, 2023) and aggregated to match the grid cell resolution using the LandInG package (Ostberg et al., 2023).
We ran the model for two forest configurations, a monoculture including only trees of Pinus sylvestris (pine monoculture hereafter) and a diverse temperate mixed forest (mixed forest hereafter). The temperate mixed forest included four major PFTs: temperate broadleaved summergreen (T-BL), temperate needle-leaved evergreen (T-NL), boreal needle-leaved evergreen (B-NL), and boreal broadleaved summergreen (B-BL). The potential trait space is defined by these four PFTs and results from all temperate and boreal needle-leaved and broadleaved trees according to the trait ranges provided in the TRY database. Via environmental and competitive filtering, however, this trait space can be smaller or change (as a result of changing environmental and demographic conditions) but still consists of different tree strategies composing the with-in PFT trait space. The simulated trait space therefore stands for the diversity of all relevant tree species. In the mixed forest configuration, any PFT can be established in any forest patch at any time following the approach used in Thonicke et al. (2020), with the establishment rate of new trees depending on light availability on the forest floor. As described above, their key functional traits are randomly sampled out of the PFT-specific ranges. These are obtained from the TRY database (Kattge et al., 2011), as described by Sakschewski et al. (2015). For the pine monoculture, only pine trees can be established, which were parameterized using the boreal needle-leaved PFT and restricting the ranges for SLA and WD to the 25th and 75th percentiles of their respective distributions in the TRY database (Kattge et al., 2011) for Pinus sylvestris. SLA and WD ranges of all PFTs and Pinus sylvestris are provided in Appendix A, Table A1. Other important differences between the tree types are their temperature limits for establishment, which reflect chilling requirements and frost tolerance, as well as their optimum temperature range for photosynthesis (see Appendix A, Table A2).
The model spinup started with the establishment of saplings on bare ground (illustrated in Billing et al., 2024, Video 1) and was run for 1000 years of simulation for each forest configuration with the spinup climate data set. Afterwards, we ran the model for 800 years with the different drought scenarios for each forest configuration (Fig. 1).
2.3 Evaluation of simulation outcomes
We evaluated the overall resilience of both pine monoculture and mixed forests against an increased frequency of extreme droughts by calculating the mean for the above- and belowground biomass (kg C m−2) across the entire study area for each year. We compared the short-term (years 1–100) and the equilibrium (long-term, years 701–800) biomass of each scenario with the baseline scenario (Scenario B). Then we analyzed different adaptation mechanisms to increased drought frequencies by calculating the mean above – and belowground biomass [kg C m−2], the mean number of trees per m2 (tree density), the mean tree height [m], the mean tree wood density [kg m−3], and the mean tree age [years] over the study region for the last 100 years of the simulation for each scenario and forest configuration. We further investigated how these mechanisms manifested in the mixed forest, including changes in the tree community composition and changes in traits for all tree individuals belonging to a particular PFT. For this, we calculate the mean tree height [m], tree wood density [kg m−3], tree age [years], SLA [m2 g−1], mortality [probability per year], and mean growth speed [years] for each PFT during the last 100 simulation years of each experiment. The growth speed of an individual tree was defined as the time to reach a height of 15 m. Trees that did not reach this size were not evaluated. For the calculation of the means for tree density, height, wood density, SLA, mortality, and age, trees were weighted with their biomass, and trees smaller than 5 m height were excluded to prevent an evaluation bias towards the multiple small trees.
Due to lack of observation data for the long-term adaptation to the occurrence of hot–dry compound events like 2018 in unmanaged temperate forests and from unmanaged temperate forests in general, no benchmarking of our model results with direct observations was possible. Instead we discuss our results qualitatively and where possible also quantitatively, referring to the findings of empirical studies from similar environments.
To assess spatial heterogeneity in forest responses, we computed the long-term (i.e., mean over the last 100 simulation years) impact of our drought scenarios on the spatial biomass variation in the pine monoculture and the mixed forest configuration across Berlin and Brandenburg.
The overall resilience in the pine monoculture and mixed forests against a higher frequency of droughts was assessed by calculating mean biomass over the entire simulation domain, assuming the Berlin–Brandenburg area is covered by vegetation only, for each simulated year and scenario (Fig. 2). While the wet scenario (Scenario A, without the 2018 drought year) and the baseline Scenario B show stable biomass in the pine monoculture forest over the entire simulation period, all drought scenarios lead to biomass loss, and biomass remains lower than under Scenarios A and B (Fig. 2a). In the mixed forest, biomass increased again after an initial phase of biomass decline of 50–150 years (Fig. 2b). Exceptions are the wet Scenario A and the most extreme climate Scenario H, where biomass declines and remains at a lower level (Fig. 2b). After an initial phase of decrease in biomass in both forest configurations, the biomass stabilized under all drought scenarios, fluctuating around a new stable state.
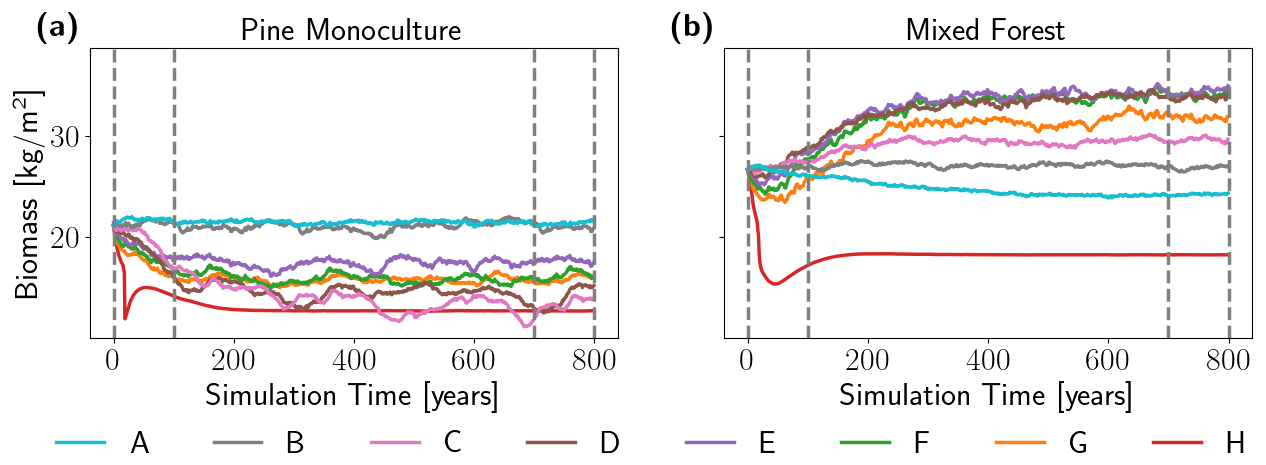
Figure 2Living biomass in pine monoculture forest (Pinus sylvestris, panel (a)) and mixed forest (b) simulated by the LPJmL-FIT DGVM and averaged over the Berlin–Brandenburg study area. Living biomass includes above- and belowground biomass [kg C m−2] and was averaged over all patches and grid cells for each year for each drought scenario (Scenarios A–H; see Table 1 for details about the scenarios). Dashed vertical lines mark the limits of the short term (ST), i.e., the first 100 simulation years, and the long term (LT), i.e., the last 100 simulation years. The wetter-than-the-baseline Scenario A and the baseline Scenario B are shown in blue and gray lines, respectively.
In the pine monoculture, the amplitude of these fluctuations varied significantly across the different scenarios, and on average, these fluctuations were much larger than those observed in mixed forests. The standard deviation of the biomass time series over the last 100 simulated years was, on average, twice as high in the monoculture (0.37 kg m−2) compared to the mixed forest (0.18 kg m−2), reflecting a stronger response to individual drought years (Fig. 2a).
Both the decrease in biomass for pine monoculture forests and the increase in long-term biomass for mixed forests were non-linear and non-monotonic; i.e, the variations in biomass levels did not linearly or monotonically correspond to the differences in drought frequencies or MCWDs among the scenarios (compare to Table 1). The lower drought frequency in Scenarios C and D resulted in a stronger decrease in biomass compared to the higher drought frequency in Scenarios E, F, and G in the pine monoculture (see Fig. 2a). In the mixed forests, Scenarios D, E, and F have a stronger increase despite a lower drought frequency than for G and H (see Fig. 2b). Under Scenarios D, E, and F, the drought frequency ranged between 0.28 and 0.53, which resulted in MCWD values of −455, −492, and −521 mm, respectively (Table 1). Surprisingly, despite the large differences in drought frequency and MCWD, the resulting biomass levels were very close under all three scenarios (Fig. 2b).
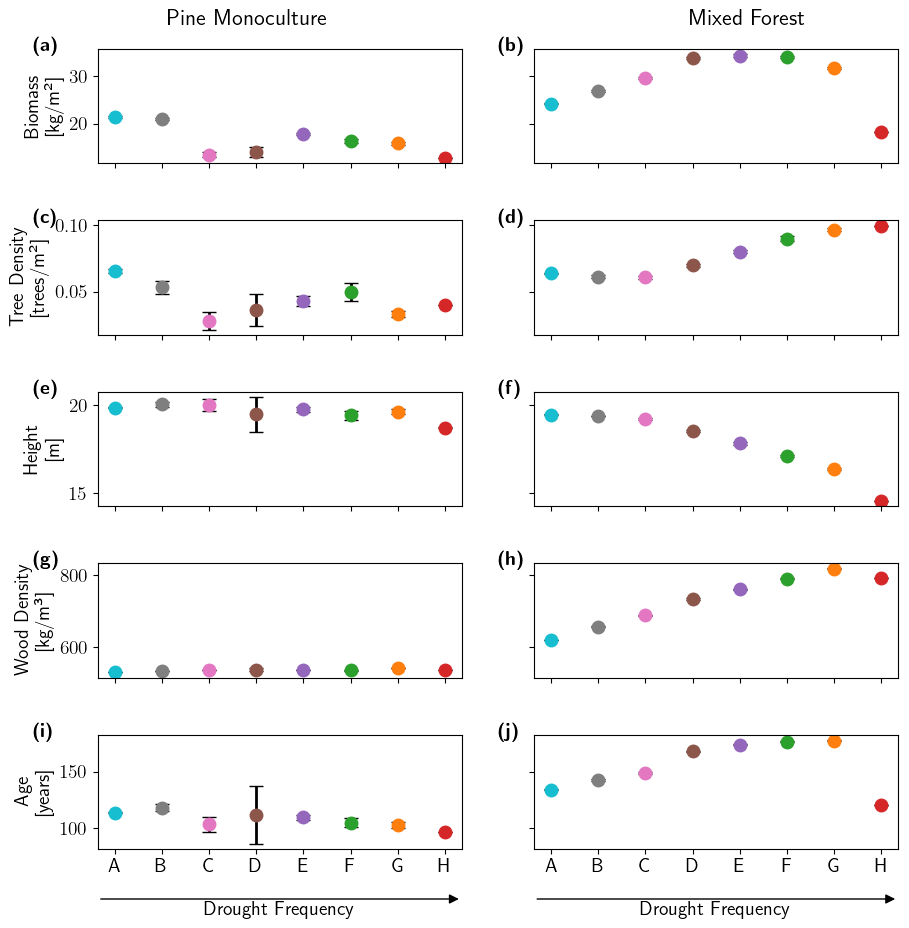
Figure 3Long-term impact of drought frequency on selected forest characteristics under Scenarios A–H as simulated by LPJmL-FIT for the pine monoculture forest (left panels) and the mixed forest (right panels) averaged over the Berlin–Brandenburg study area. Biomass (a, b), tree density (c, d), height (e, f), wood density (g, h), and age (i, j) are displayed as means over the last simulated 100 years (simulation years 701–800).
The long-term responses of monocultures and mixed forests to increased drought frequency differed at both the community level and the level of individual trees (Fig. 3). At the community level, monoculture vs. mixed forests showed opposing responses towards increased drought frequency. In the pine monoculture forest, long-term biomass (Fig. 3a) and tree density (Fig. 3c) were higher in Scenarios A and B compared to all the drier scenarios (Scenarios C–H). The mixed forest showed a different pattern. Here, biomass (Fig. 3b) and tree density (Fig. 3d) were higher at the end of the simulation period the higher the drought frequency became. However, under the extreme Scenario H biomass was lower than under Scenarios A and B, while the number of trees was highest (Fig. 3b and d). While height and wood density showed little variation or no trend across the scenarios for the pine trees growing in the monoculture forest (Fig. 3e and g), increasing drought frequency in the mixed forest led to decreasing tree height (Fig. 3f) and increased wood density (except under Scenario H; see Fig. 3h). Mean forest age was lower under Scenarios C–H in the pine monoculture forest compared to Scenarios A and B and showed little variation (Fig. 3i). In contrast, trees in the mixed forest grew older the higher the drought frequency became, again with the exception of Scenario H, where average tree age was approx. 50 years lower (Fig. 3j). In general, there was much less adaptation in individual tree properties and total stand properties in the monocultures compared to the mixed forests. In mixed forests, trees became smaller, had a higher wood density, and grew older with increasing drought frequency, while the monocultures did not show clear trends in the properties of individual trees. It seems that tree demography effects in conjunction with trait adaptation at the individual level dominate forest adaptation that resulted in the hump-shaped biomass pattern (Fig. 3b). The differences between the performance of pine monocultures and mixed forests under increasing drought frequencies were also confirmed by the spatial pattern of simulated long-term biomass in the Berlin–Brandenburg study area (see Appendix B, Fig. B1). A new aspect was, however, that we found large spatial heterogeneity for the performance of both the pine monoculture and the mixed forest under baseline climate (Scenario B, Fig. B1a, d). Pine monocultures seemed especially heat-sensitive, as Berlin and urban areas of Brandenburg, which have a higher temperature (see Appendix B, Fig. B2), showed much less biomass than in rural areas (Fig. B6a–c). In contrast, mixed forests seemed to respond to both temperature increase and water deficits (see Appendix, Fig. B3).
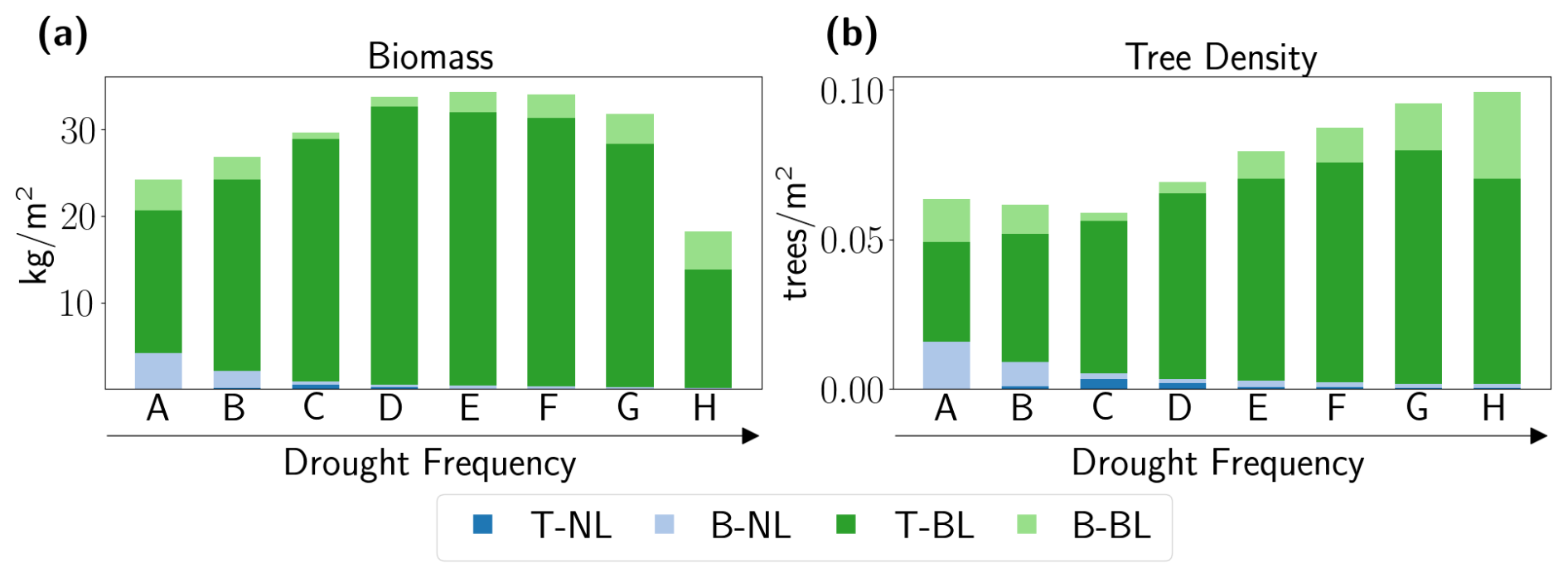
Figure 4Composition of the plant community in the mixed forests averaged over the Berlin–Brandenburg simulation domain and the last 100 simulation years (701–800). Biomass [kg m−2] (a) and tree density [trees m−2] (b) of tree individuals belonging to temperate needle-leaved evergreen PFT (T-NL, dark blue), boreal needle-leaved evergreen PFT (B-NL, light blue), temperate broadleaved summergreen PFT (T-BL, dark green), and boreal broadleaved summergreen PFT (B-BL, light green) for each drought frequency scenario.
In addition to changes in biomass and tree density in the mixed forests (as seen in Fig. 3), their functional composition also shifted in response to increasing drought frequency (Fig. 4). The higher the drought frequency, the lower the proportion of needle-leaved trees belonging to the T-NL and B-NL PFTs was. Even though needle-leaved trees could still adapt under Scenarios D and E, their contribution to the overall biomass was marginal in the equilibrium state. Generally, broadleaved trees dominated the forest community with their biomass being 12 times as high as needle-leaved trees, and their tree density became 6 times as high as in the baseline Scenario B. While the biomass and tree number of needle-leaved trees further declined with increasing drought frequency, the number of broadleaved trees increased, and their total biomass until drought frequency was also too high (Scenarios F–H). That is, the patterns observed in Fig. 3 can be mainly attributed to the increasing dominance of broadleaved trees. In particular, the temperate broadleaved tree is the dominant PFT in all scenarios, with its highest fraction in intermediate drought scenarios (Scenarios C–E) and its lowest fraction in the wettest scenario. In contrast, the fraction of the boreal broadleaved PFT is highest in the wettest and driest scenarios, and the boreal needle-leaved PFT plays only a minor role in the composition, with its highest fraction in Scenario C. Interestingly, the tree density of boreal broadleaved trees (B-BL) is small under Scenarios A and B, the lowest under Scenario C, and increasingly higher under Scenarios D–H (Fig. 4b) but does not contribute the same proportion of biomass (Fig. 4a). Given those patterns found, it seems that the tree individuals in each PFT adapt differently to the new climate normals.
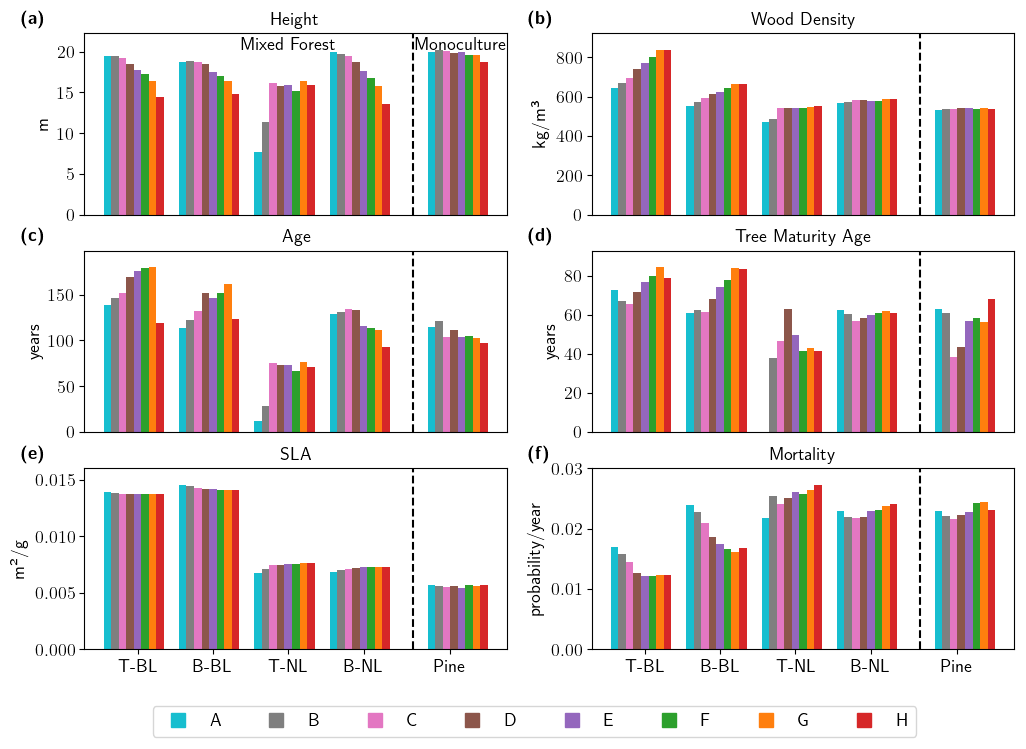
Figure 5Long-term impact of drought frequency on selected tree characteristics. Biomass-weighted mean values are shown for (a) height [m], (b) wood density [kg m−3], (c) age [years], (d) tree maturity age [years] (also referred to as growth speed, defined as tree height at which a tree reached a height of 15 m), (e) specific leaf area [m2 g−1] (SLA), and (f) mortality [probability per year] at the end of the simulation period (simulation years 701–800) in pine monoculture forest (pine) and for each PFT in mixed forests under drought frequency Scenarios A–H. Temperate broadleaved trees (T-BL), boreal broadleaved trees (B-BL), temperate needle-leaved trees (T-NL), and boreal needle-leaved trees (B-NL). Please note that values for T-NL are based on very low numbers of individuals (see Fig. 4b) and that the sample size for the calculation of tree maturity age is less compared to the data in the other panels, as not all trees reach a height of 15 m.
Figure 4 showed that changes in biomass and tree numbers could mainly be explained by shifts in the functional composition of mixed forests, i.e., how much biomass and how many trees belonging to a particular PFT contributed to the forest community. In addition, we observed drought-induced changes in the characteristics of individual trees across PFTs concerning their height, wood density, and age (Fig. 3e–j). In Fig. 5, we can show that these changes were attributed to trait adaptation within each PFT, leading to shifts in mean characteristics of each PFT, rather than to shifts in PFT dominance. The trends in drought-induced shifts were mostly similar between PFTs for height and wood density but differed for the age of individual trees. Except for temperate needle-leaved trees (which are very low in number and therefore need to be treated with care), the mean height of all PFTs decreased from around 20 m to around 15 m in mixed forests (Fig. 5a). Pine trees growing in a monoculture only slightly decreased in height; i.e., intra-species plant competition seems to strongly impact drought adaptation. For wood density (Fig. 5b) we observed differences between broadleaved species, which strongly increased in density (by 29.6 % for T-BL and 19.8 % for B-BL), and needle-leaved species, which initially started with a lower wood density and showed only marginal increases from Scenarios C to H. Pine trees in monocultures showed even less response than the boreal needle-leaved type. For mean tree age, broadleaved trees generally grew older, while at least the boreal needle-leaved PFT showed a younger age structure with increasing drought. Again, the results of the temperate needle-leaved PFT need to be treated with care because of low numbers. In monocultures, the mean age of pine trees was slightly lower than of their pendant in the mixed forest. Growth speed was generally faster for needle-leaved trees (40–65 years to reach a size of 15 m) than for broadleaved trees (60–85 years). While boreal and temperate broadleaved trees have a similar growth speed despite their differing wood density, boreal needle-leaved trees seem to grow more slowly than temperate ones. In most cases, a higher drought frequency slows down the growth speed; i.e., trees need longer to reach a height of 15 m. However, pine trees in monocultures seem to reach this height faster under mild drought scenarios (Scenarios C and D) and similarly to the baseline scenario for more frequent droughts (Scenarios E–H). For broadleaved trees mortality decreased with increasing drought frequency, and in all drier-than-baseline scenarios (Scenarios C–H) it was lower than for the needle-leaved trees (Fig. 5f). In contrast, for needle-leaved trees the mortality for the driest scenarios (Scenarios D–H) was higher than for the baseline scenario. Also for SLA, broadleaved and needle-leaved trees showed opposing trends, although changes were relatively small for all tree types (Fig. 5e). For the needle-leaved trees SLA increased with drought frequency, while for the broadleaved trees SLA decreased slightly, and for pine there was no significant trend (Fig. 5e). For broadleaved trees SLA was higher than for needle-leaved trees, for which in turn SLA was higher than for pine trees and even exceeded the upper limit of the Pinus sylvestris parametrization range.
The spatial pattern of simulated long-term biomass in the Berlin–Brandenburg study area confirmed the differences between the performance of pine monocultures and mixed forests under increasing drought frequencies (Appendix B, Fig. B1). A new aspect was, however, that we saw large spatial heterogeneity for the performance of both the pine monoculture and the mixed forest under baseline climate (Appendix B, Fig. B1a, d). Pine monocultures seemed especially heat-sensitive, as Berlin and urban areas of Brandenburg, which have a higher temperature (Appendix B, Fig. B2), showed much less biomass than in rural areas (Appendix B, Fig. B1a–c). In contrast, mixed forests seemed to respond to both temperature increase and water deficits (see Appendix, Fig. B3). Under increased drought frequency (Scenarios E and H shown here), areas of very low biomass in pine monocultures became larger. In the extreme Scenario H, they basically collapsed in the western–central Berlin–Brandenburg area and near the river Elbe in northwest Brandenburg (Appendix B, Fig. B1c). As shown before, an increase in drought frequency led to an overall increase in biomass of mixed forests if drought was not occurring every year (Scenario H, Appendix B, Fig. B1e). As a result, biomass in the western–central part of Brandenburg but not in Berlin was higher in Scenario E but drastically lower in Scenario H (Appendix B, Fig. B1e, f).
Summarizing all of the above results, we observed that mixed forests could better adapt to increasing drought frequency than pine monoculture forests. Mixed forests can adapt via shifts in PFT composition as well as functional and structural trait shift at the community level resulting from changes in the characteristics of the tree individuals: the fraction of broadleaved trees increased, and these became sturdier (smaller with higher wood density) and grew older. However there were spatial differences throughout the study areas, with Pinus monocultures especially suffering along urban areas and mixed forests responding stronger in the western–central part.
4.1 Differences between pine monocultures and mixed forests under current conditions
Generally, our results for pine monocultures show lower total biomass (around 21 kg m−2) under present conditions (baseline scenario) than mixed forests (around 27 kg m−2). This is in line with a study across forests in northern Germany (Förster et al., 2021) that compared landscape-scale aboveground biomass of different forest types and found much lower values for old-grown pine forests (19 kg m−2) than for forests with natural development (32 kg m−2). Moreover, multiple forest studies show a positive productivity–biodiversity relationship and higher carbon stocks in more diverse forests (Watson et al., 2018; Ali and Yan, 2017; Zhang and Chen, 2015), which can, however, strongly depend on species identity (Chisholm and Dutta Gupta, 2023) and climatic conditions. In a US-scale analysis of forests, Fei et al. (2018) found a positive biodiversity–productivity relationship in arid to semi-arid areas, which turned negative under a more humid climate.
Also the difference in mean tree age between monocultures (118 years) and mixed forests (143 years) is similar to observed values from Förster et al. (2021), who found a mean stand age of 112 years for old-grown pine forests and 146 years for naturally developed forests. However, our findings on stem density, which show a slightly higher number of trees per area in mixed forests, deviate from their results, as they find a much higher number of trees in young and old pine forests than in mixed forests with natural development. This can be an effect of management rather than of tree species composition. However, other studies confirm our results, with higher tree numbers per area in mixed forests than in pine monocultures, e.g., due to a higher use efficiency of light, water, or nutrients by species mixing (Pretzsch and Biber, 2016; Pretzsch and Schütze, 2016). Moreover, our results for slightly higher tree heights and faster growth speed in monocultures are confirmed by other studies (Pretzsch and Forrester, 2017), which is one of the reasons why pine forests have been promoted in forestry in the past decades (Spathelf and Ammer, 2015).
4.2 Effects of droughts on biomass and structure of pine monocultures versus mixed forests
Under drier and warmer conditions the difference in total biomass between pine monocultures and mixed forests increases strongly, as the biomass of the monocultures declines further, while the biomass of mixed forests increases in the long-term. The stronger fluctuations in the biomass time series also show that the pine forest responded stronger to individual drought years, which indicates increased system instability. This is in accordance with observations, which found that pine monocultures were affected by the European drought year 2018, which led to high mortality rates of Pinus sylvestris and a vegetation shift to other species (Haberstroh et al., 2022). In addition to the drier conditions, one major reason for the biomass decline in the pine monoculture was the warmer temperatures negatively affecting the establishment of pine trees (compare Appendix B, Fig. B1a–c with Appendix A, Table A2), which germinate better with cold stratification (Houšková et al., 2021) and have a lower thermostability of photosystem II during photosynthesis compared to other important temperate tree species (Húdoková et al., 2022). However, the model might estimate the chilling temperature needed for cold stratification to be too low for the B-NL PFT and pine trees in Brandenburg, as germination requirements vary with provenance (Hannerz et al., 2003; Rampart, 2018; Tillman-Sutela and Kauppi, 1995) and might be different for temperate lowlands like Brandenburg compared to colder regions. It is surprising that despite the lower drought frequency in Scenarios C and D, simulated biomass is lower than under scenarios with higher drought frequency (Scenarios E–H). This can, however, be explained by changes in the seasonal precipitation distribution, where a rather wet April and March in 2018 favored tree productivity before the hot–dry conditions occurred in summer and started to stress the trees (Appendix D, Fig. D1). A low 2018-like frequency means more average growing conditions in early spring followed by a hot–dry summer, which still reduces biomass. A higher frequency of 2018-like drought conditions means above-average growing conditions in early spring but more severe drought conditions in summer, which results in a pine monoculture of slightly smaller and younger trees storing more biomass (see Fig. 5 for details).
Broadleaved trees, e.g., European beech Fagus sylvatica or even pedunculate oak Quercus robur, also strongly suffered from the 2018 drought, but these were less strong, and the mixing of appropriate species might have been able to mitigate drought impacts (Schuldt et al., 2020). In our simulations, we also see that the biomass of mixed forests drops in the first decades, as they need time to adjust to altered climate conditions. However, after this phase of adjustment, biomass in mixed forests increases in the long term (except for the extreme Scenario H). As comparative data on such long-term forest dynamics are difficult to obtain, it is helpful to assess the validity of our results with the help of a spatial aridity gradient. A comprehensive study on data from a tree ring database along an aridity gradient in Europe (Pardos et al., 2021) tested the growth response to site-specific drought events between 1975 and 2015 for monospecific and mixed stands. They generally found a higher resilience and resistance to drought events in mixed forests compared to monocultures but especially an increased resilience to drought at drier sites, with slight increases in productivity in post-drought years for mixed forests but not for monocultures. Also a long-term study using inventory data of boreal forests of western Canada from 1958 to 2011 confirmed that with increasing temperature and decreasing water availability, biomass declined in species-poor forests, while it increased in species-rich forests (Hisano et al., 2019). These two studies might be an indicator that well-adapted species compositions might indeed become more productive and increase their biomass over centuries in mixed forests. However, the increased biomass simulated by LPJmL-FIT might be overestimated because the model did not include nutrient limitation, e.g., nitrogen, which might limit tree growth to a greater extent under drier conditions.
A higher drought frequency impacts not only the biomass of forests, but also their structure. We found large differences between the pine monoculture and a mixed forest, where different properties of the mixed forest responded much more strongly to increased drought frequency. In the pine forest, tree density decreased in response to more droughts and higher temperatures, which was also found by Haberstroh et al. (2022). Surprisingly, the pine monoculture does not exhibit the expected shift towards higher wood density under drought conditions. One explanation lies in the inherent structure of the Pinus stands, which contain a dense population of fast-growing trees with lower wood density under present conditions. These trees, characterized by rather fast vertical growth, may outcompete trees with higher wood densities due to light competition. Therefore, the presence of trees with lower wood densities could hinder the growth success of more slowly growing trees, although trees with higher wood densities might be better adapted to drought conditions. This competition effect could lead to lower drought resistance of pines growing in a monoculture compared to pines in more diverse forests, as also found in an empirical study (Granda et al., 2018). Such a phenomenon suggests that light competition in the understory plays a significant role in developing certain drought-adapted plant strategies. Similar shifts in wood densities as a response to increased competition have been previously observed in earlier studies (Billing et al., 2024). As the mean height and age only decreased slightly, the decrease in pine forest biomass seems to be especially caused by lower tree densities and not by changes in tree morphology.
In contrast, tree density increased in mixed forests, even for the most extreme drought Scenario H. Individual trees, however, had higher wood density, leading to slower growth, reduced mortality, smaller SLA, and smaller height while getting older. Higher wood density with decreasing annual rainfall (below values of 1000 mm a−1, which is the case in our study area) was also found in a global tree inventory analysis for temperate forests (Bouchard et al., 2024). But also species-specific investigations showed increasing wood densities for the same species from different provenances across the world with higher aridity (Nabais et al., 2018). Fei et al. (2017) also observed that in the eastern part of the US, tree species whose range shifted to drier areas had higher median wood density. When trees invest more carbon into their wood density, this can lead to a decrease in growth rate and size (Aiba and Nakashizuka, 2009; Kallarackal and Ramírez, 2024). Moreover, it makes sense that trees optimize their height towards smaller sizes, as found in our study, as large trees suffer more strongly from droughts (Bennett et al., 2015). Ryan and Yoder (1997) also found that for the same species, its maximal height can differ strongly between locations, with smaller tree sizes found in more arid environments. More slowly growing forests also have a lower turnover rate (Black et al., 2008; Johnson and Abrams, 2009), which is reflected in our finding of an older mean tree age under higher drought frequency (except for the most extreme Scenario H).
4.3 Underlying mechanisms leading to a higher resilience of the mixed forest
As discussed in the previous section, mixed forests seemed to have a higher resilience towards droughts: their biomass stabilized at high values after an initial adaptation phase, and trees had a higher wood density and grew more slowly but got older and reached a lower height. Here, we discuss (i) if the higher resilience is rather a result of a shift in the composition of the community or in the traits of individual PFTs towards more optimized values, as well as (ii) the general role of species diversity in forest resilience.
Our results showed a shift in PFT composition in the mixed forest: while needle-leaved trees declined strongly in their biomass and tree density, playing only a marginal role under increased drought, the biomass and tree density of broadleaved trees increased. These higher tree densities with increasing aridity have also been observed in a study in northern Germany on Fagus sylvatica monocultural stands (Weigel et al., 2023). Such a community shift from pine trees to broadleaved trees in response to the drought in 2018 has also been observed in satellite data and tree mortality data in Germany (Haberstroh et al., 2022). Particularly, temperate broadleaved trees overall benefited from an increased frequency of the drought year 2018, as they are less limited by higher temperatures (Appendix A, Table A2). Looking back at our question if the shift in the plant community composition or a shift in individual tree traits increases the forest resilience of mixed forests, we can say at this point that a shift towards more temperate broadleaved trees (and the associated shift in community-weighted mean traits) can at least partly explain the higher resilience of mixed forests.
The trait shift in individual PFTs as a response to a higher drought frequency, however, seems much more important. The flexible-trait scheme of our model allows the emergence of different plant strategies to optimize plant performance under stressful conditions within a PFT. That is, from our model results, we can learn about PFT-specific plant trait combinations that are best adapted under different drought conditions. In contrast to needle-leaved species, it seems to be optimal for broadleaved species to strongly invest in wood density under higher drought frequency. This resembles the well-known coping mechanisms towards high wood densities and slightly smaller SLAs under drought or dry summer conditions observed at broader scales (Greenwood et al., 2017; Serra-Maluquer et al., 2022) and explains the decrease in mortality (Greenwood et al., 2017) for broadleaved trees and the overall increase in wood density at the community level. In the study on Fagus sylvatica monoculture stands in Germany, lower growth was observed in response to a decadal decrease in the climatic water balance (Weigel et al., 2023). In general, there seems to be a shift for both broadleaved PFTs towards a more conservative strategy, where they invest in wood density and grow more slowly (Chave et al., 2009; Poorter et al., 2010) and less tall (Aiba and Nakashizuka, 2009; Kallarackal and Ramírez, 2024) but become older (Laurance et al., 2004; Black, Colbert, and Pederson 2008; Bigler and Veblen, 2009). In contrast, boreal needle-leaved trees are less productive and have a lower wood density and thus a higher mortality, and consequently the forests are composed of younger and smaller individual trees with few trait adaptations, which we also see for pine monocultures. Note that the smaller trait ranges from which possible SLA and WD values are drawn for pine trees and needle-leaved PFTs compared to the ranges for broadleaved PFTs by design result in fewer possible trait combinations and therefore fewer possible ways to adapt. However, pine trees do not and the needle-leaved PFTs only slightly adapt via increased WD and do not use the still open niche space. The long-term mean WD of the pine trees and needle-leaved PFTs remains well below the upper limit defined in the parametrization of LPJmL-FIT (see Fig. 5b and Appendix A, Table A1). In the case of SLA of pine trees and needle-leaved PFTs the mean of its SLA distribution is rather close to the upper parametrization limit in all scenarios and is closer to the limit with increasing drought frequency (see Fig. 5e and Appendix A, Table A1). Figure C1 exemplarily shows for Scenarios B and H that the SLA distribution for the pine trees and needle-leaved PFTs is cut off at the upper limit. This means that with increasing drought frequency the environmental and competitive filtering would most likely result in pine trees and needle-leaved PFTs with SLA values higher than those allowed by parametrization and closer to those of the broadleaved trees if there were no limits set. That shows that according to our experiments needle-leaved trees with WD and SLA values in the range of the observations collected in the TRY database for temperate and boreal needle-leaved species do not perform well under scenarios with higher drought frequency.
In general, mixed-species forests have been discussed as an adaptation strategy to reduce the risk faced by forest ecosystems under global change (Forrester et al., 2016). One reason is the potential niche complementary of different species, reducing competition for resources (Morin et al., 2011) and improving the resource supply and uptake (Richards et al., 2010). Moreover, interspecific facilitation can partly release trees from stress, leading to higher resistance and resilience of mixed forests, especially to climate extremes such as droughts (Pretzsch et al., 2013). However, these findings can strongly depend on species identity and the environmental context (Decarsin et al., 2024; Forrester et al., 2016). Therefore, the forest species mixture has to be appropriately chosen for a specific stand to increase the likelihood that beneficiary effects mitigate drought impacts (Ammer, 2017). As we do not account for these facilitative effects in our simulations, we might even underestimate the positive effects of mixed forests on drought resilience in our assessments.
In conclusion, it can be summarized that numerous mechanisms lead to the higher resilience via adaptation of mixed forests to an increased drought frequency, which we have only partially considered in this study. The ability of mixed forests to adapt to increasing drought frequency stems from establishing new, especially broadleaved, trees whose trait combinations have a higher drought tolerance, which results in a trait shift. However, the observation that biomass decreased again when drought frequency was too high shows that in a mixed forest, the adaptation capacity also has limits beyond which productivity decreases.
4.4 Implications
We found that an increased drought frequency along with increased mean temperatures adversely affects the productivity of forests in Berlin–Brandenburg in the first decades. This leads to a massive biomass decline in both forest types, pine monocultures as well as mixed forests, with multiple implications for ecosystem functions and services (for examples see Brockerhoff et al., 2017). However, we also saw that mixed forests can adapt in the long term (as long as extreme droughts do not occur too often) by a change in PFT composition towards more broadleaved trees, but also by shifts in PFT traits. Such a shift towards a higher fraction of broadleaved trees was also suggested in a Europe-wide study combining forest inventory data with climate data driven by different representative concentration pathway (RCP) scenarios, which found a reduction in Pinus sylvestris and Picea abies (Norway spruce) from lowlands in Central Europe to higher altitudes or more northern areas, but extended areas for Quercus robur (pedunculate oak) and ambivalent results for Fagus sylvatica (Buras and Menzel, 2019). In reality, various factors can prevent plant species from occupying all areas that meet their environmental requirements (Lehsten et al., 2019; Lenoir and Svenning, 2015; Thompson and Fronhofer, 2019; Zani et al., 2023), which also depends on the dispersal mechanism, such as seed versus pollen dispersal (Austerlitz and Garnier-Géré, 2003; Cheng et al., 2024; Gerber et al., 2014; Kim et al., 2022). In our model, we do not simulate different dispersal strategies. PFTs and trait combinations become established everywhere with the same probability. That means there is no trait inheritance, and therefore the trait combination of new saplings is independent of the previous population's trait distribution. Therefore, especially with respect to trees with local seed dispersal, our simulations might underestimate the time needed for trait shifts and changes in PFT compositions to happen without human intervention. Another limitation of our methodology is the lack of observation data regarding the long-term adaptation of temperate natural forests to increased frequency of extreme hot–dry compound events and therefore the lack of benchmarking of our model results with direct observations.
The current and potential future impacts of climate change have concerned practitioners and scientists for more than 2 decades (Hanewinkel et al., 2022). However, a case study with forest practitioners in four regions of Germany by Milad et al. (2013) showed that strategies for adapting forest management were at that point still in their early stages. Recently, the Scientific Advisory Board for Forest Policy of the German Federal Ministry of Food and Agriculture proposed mixed forests to better cope with climate change, but also the active introduction of better-adapted tree species that are taxonomically, spatially, and ecophysiologically closely related to current species to also support native biodiversity (Bauhus et al., 2021). Moreover, using seeds from seed provenances adapted to future climates for reseeding rather than local seed provenances might further mitigate the initial biomass declines that we found in our simulations, as a recent study on assisted tree migration in Europe showed (Chakraborty et al., 2024). In order to apply these suggested methods for increasing the resilience towards possible new climate normals, knowing which trait combinations and species communities perform best under these conditions is crucial. With our study, we contribute to this quest by showing which forest structure and tree characteristics result under increased drought frequencies in unmanaged forests in Berlin and Brandenburg.
Due to the small grid size of only 2 × 2 km, we were able to observe large heterogeneity in the forest biomass across Berlin–Brandenburg. This heterogeneity increased under increased drought frequencies. However, this could also be a consequence of the specific spatial heterogeneity in the climate of the year 2018, which has an increasing influence on the results the higher the frequency becomes. The observed heterogeneity underscores that for the management of forests, site-specific solutions are required that account for things like temperature differences between more urban and more rural areas, the rainfall gradient in Berlin–Brandenburg, and different soil textures. Furthermore, they highlight that in addition to local studies there is a strong need for high-resolution climate projections that accurately reflect possible increases in extreme drought frequencies and models that accurately simulate the impacts of these climate projections on vegetation.
Our results suggest that increased drought frequencies could lead to a reduction in biomass in both pine monoculture forests and mixed forests in Brandenburg and Berlin. Mixed forests, however, might eventually recover and even exceed initial biomass levels in the long-term, as long as drought frequencies are not too high. The higher resilience of mixed forests in our simulations was due to higher adaptive capacity. The adaptation, however, profoundly changed forest characteristics: mixed forests were predominantly composed of smaller, broadleaved trees with higher wood density and slower growth, which can be summarized as a shift towards more conservative strategies. These changes would have significant implications for forestry, even when sustainably managed; related industries; and other ecosystem functions and services. Our results thus highlight the importance of incorporating biodiversity into forest management and preparing for shifts in the ecosystem services provided by forests in Brandenburg and Berlin in the future.
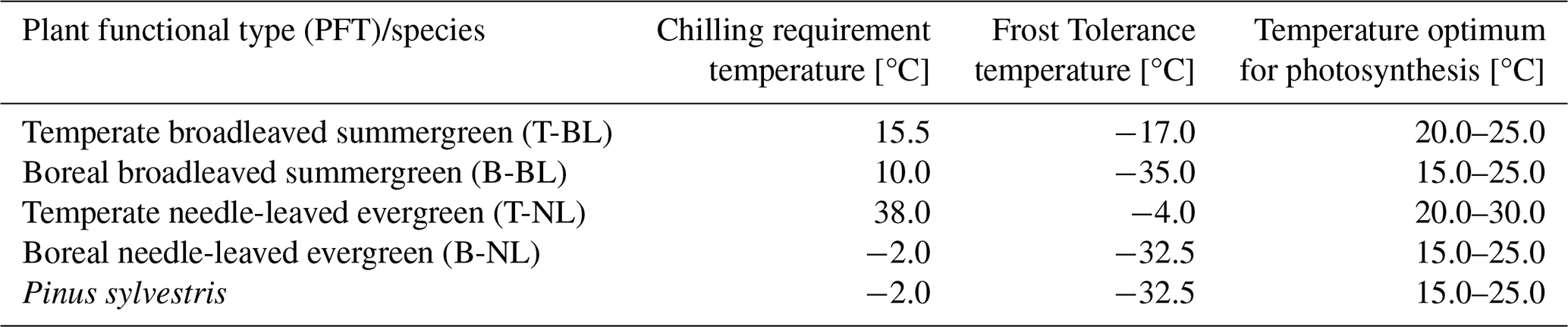
Table A1Specific leaf area [m2 g−1] (SLA) and wood density [kg m−3] (WD) ranges for the simulated plant functional types and Pinus sylvestris.
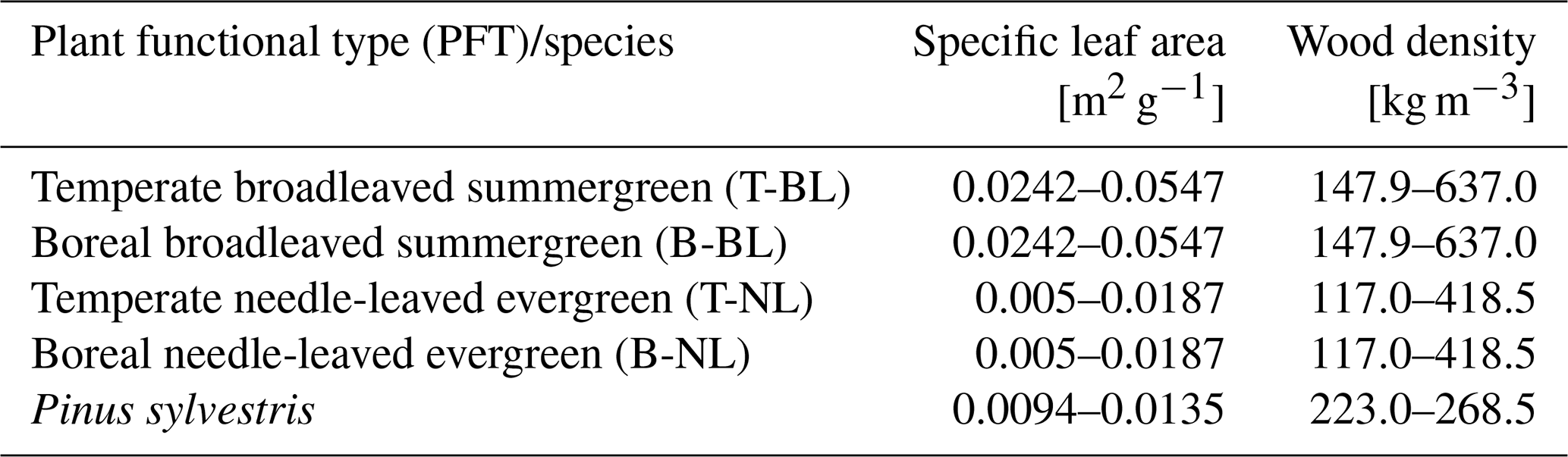
Table A2Temperature limits for tree establishment and survival as well as optimum temperature range for photosynthesis. For establishment to happen, the mean of the annual minimum temperature over the last 20 years must be larger than or equal to the frost tolerance temperature [°C] and smaller than or equal to the chilling requirement temperature [°C]. Trees die if the mean of the annual minimum temperature over the last 20 years is smaller than the frost tolerance temperature [°C]. The temperature optimum for photosynthesis is the temperature range in which photosynthesis is not inhibited by too low or too high temperatures.
Under the baseline scenario, the more or less temporarily stable biomass levels (Fig. 2b) result from biomass values of around 30 kg m−2 in southern Brandenburg and slightly lower biomass values of around 20 kg m−2 in northern parts (Fig. B1d). The higher biomass under medium-frequency Scenario E compared to the baseline (Fig. 2b), which is driven by changes in structural and functional traits (Figs. 3 and 4), is mainly found in central–western and southern parts and also in isolated areas in eastern parts of Brandenburg, where biomass reaches values of around 40 kg m−2 (Fig. B1e). When the 2018 drought becomes the new climate normal (Scenario H), lower biomass values (< 20 kg m−2) show little spatial variation across the study area, with slightly lower values in western Brandenburg.
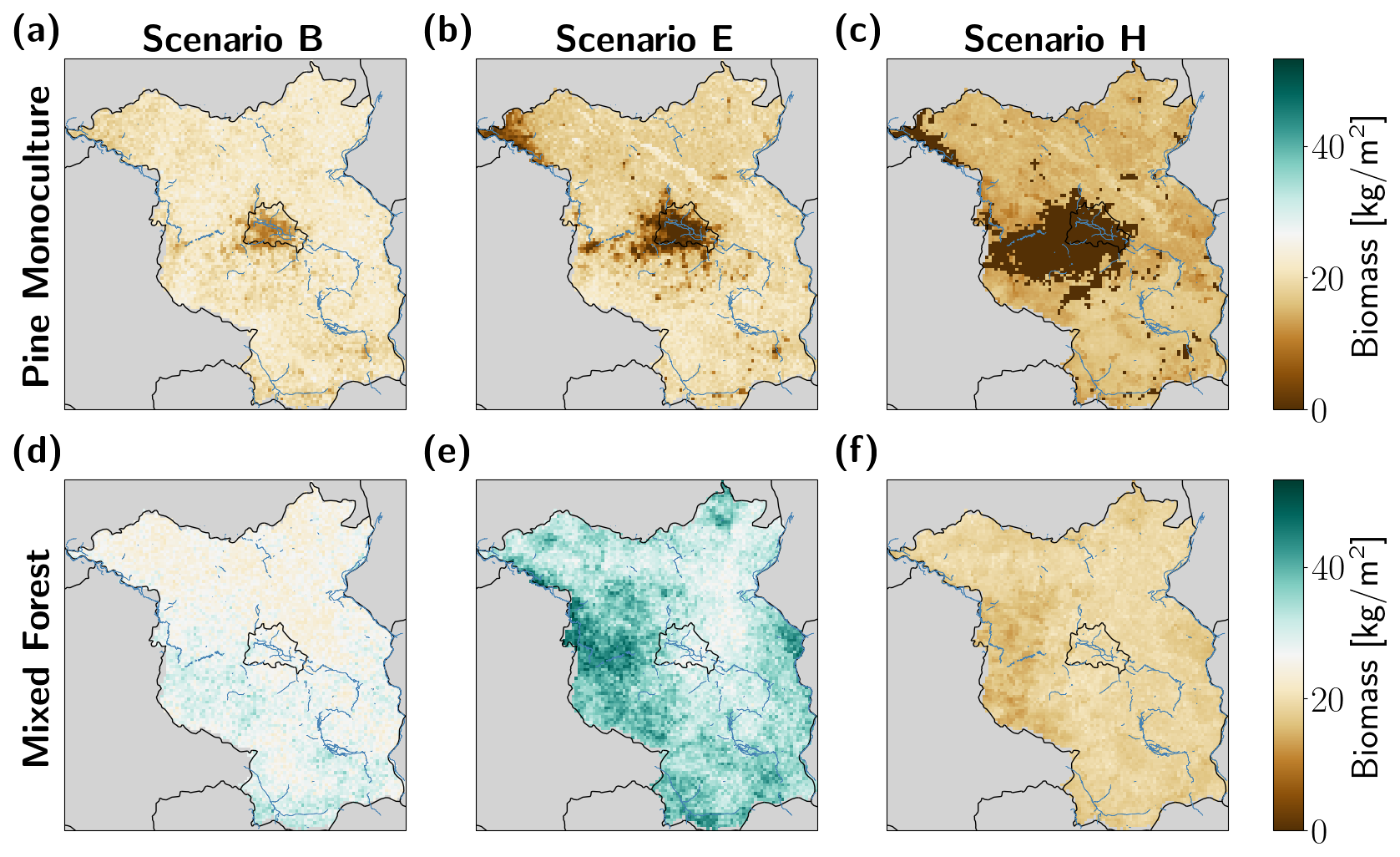
Figure B1Simulated long-term (mean over last 100 simulation years) biomass under selected drought frequency scenarios (baseline (Scenario B), example for medium-frequency scenario (Scenario E), and highest drought frequency (Scenario H)) for pine monocultures (top row) and mixed forests (bottom row) for the Berlin–Brandenburg study area. The state borders of Berlin and Brandenburg are shown in black, major riverbanks in blue. Biomass in pine monoculture decreased under all scenarios (a–c), especially in central and western parts, whereas spatial patterns of biomass increased differently in mixed forests under each scenario (d–f). See Appendix B, Fig. B2 for Scenarios A, C, D, F, and G.
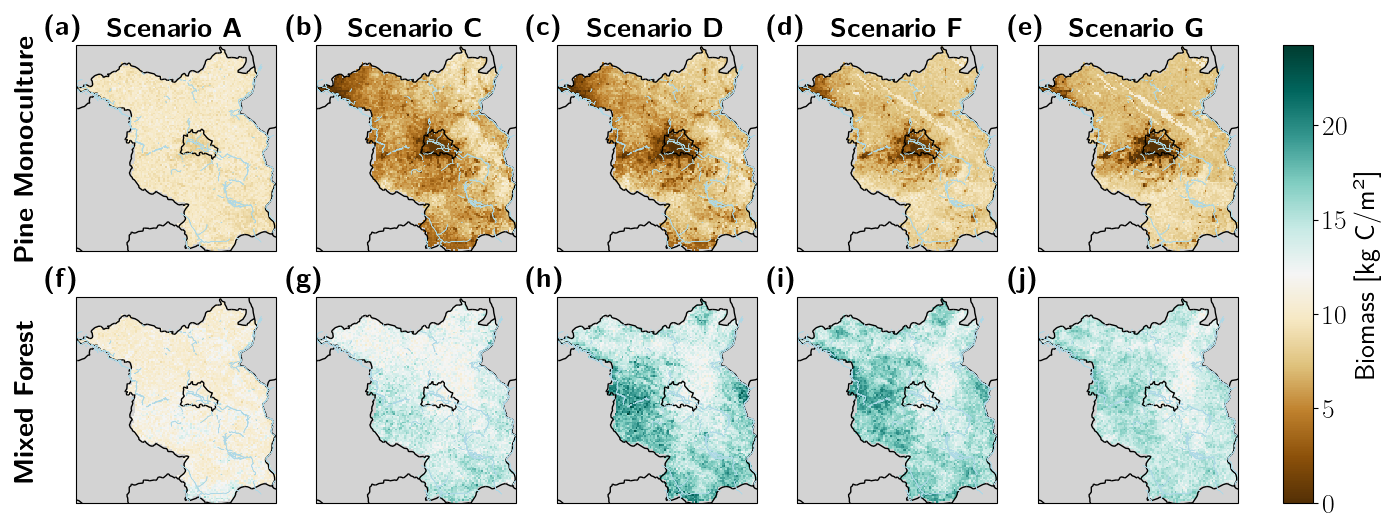
Figure B2Simulated long-term (mean over last 100 simulation years) biomass under selected drought frequency Scenarios A, C, D, F, and G for pine monocultures (a–e) and mixed forests (f–j) for the Berlin–Brandenburg study area. The state borders of Berlin and Brandenburg are shown in black, major river banks in blue.
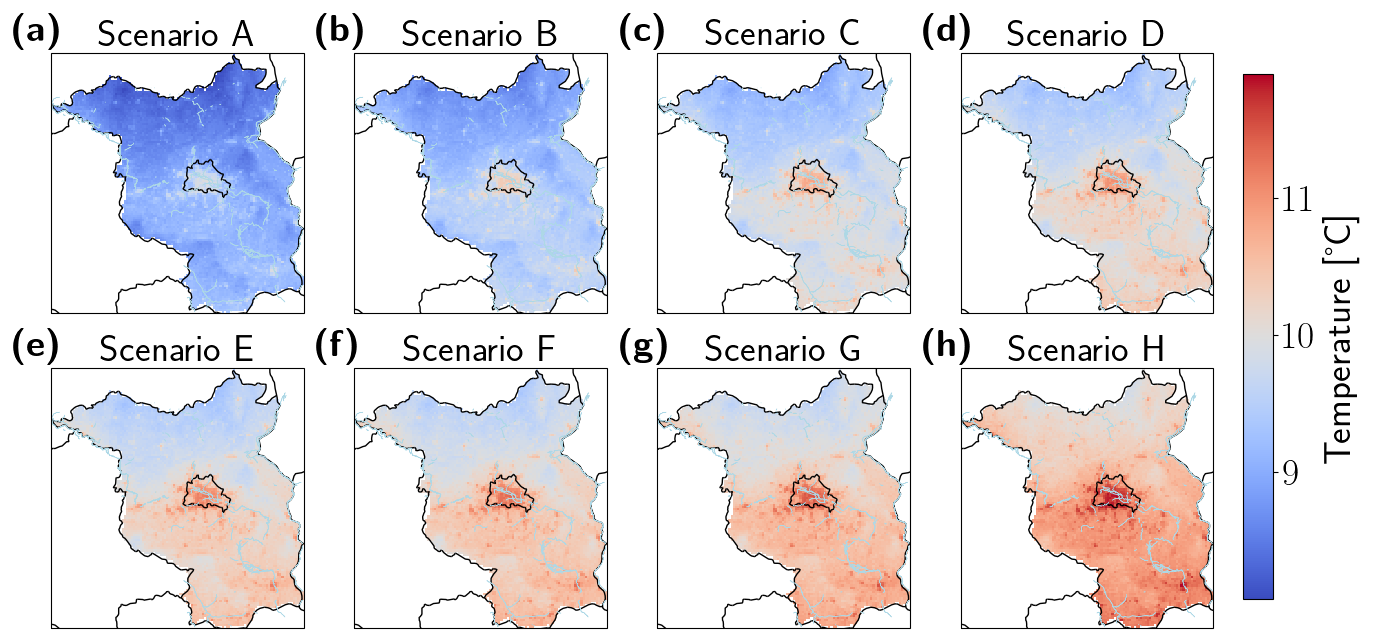
Figure B3Mean daily temperatures [°C] over all simulation years for the Berlin–Brandenburg area for scenarios with increasing frequency of the year 2018 from A–H (a–h). The state borders of Berlin and Brandenburg are shown in black, major river banks in blue.
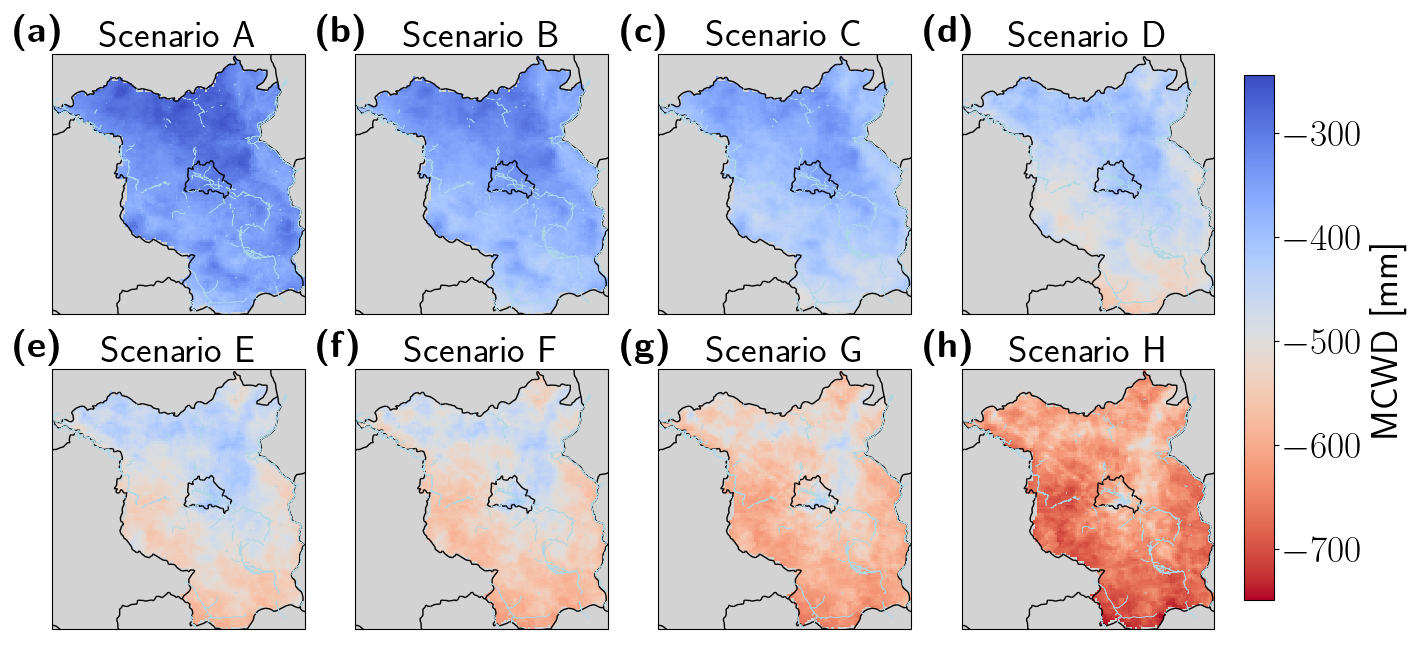
Figure B4Mean maximum annual climatic water deficit (MCWD) [mm] over all simulation years for the Berlin–Brandenburg area for scenarios with increasing frequency of the year 2018 from A–H (a–h). The state borders of Berlin and Brandenburg are shown in black, major river banks in blue.
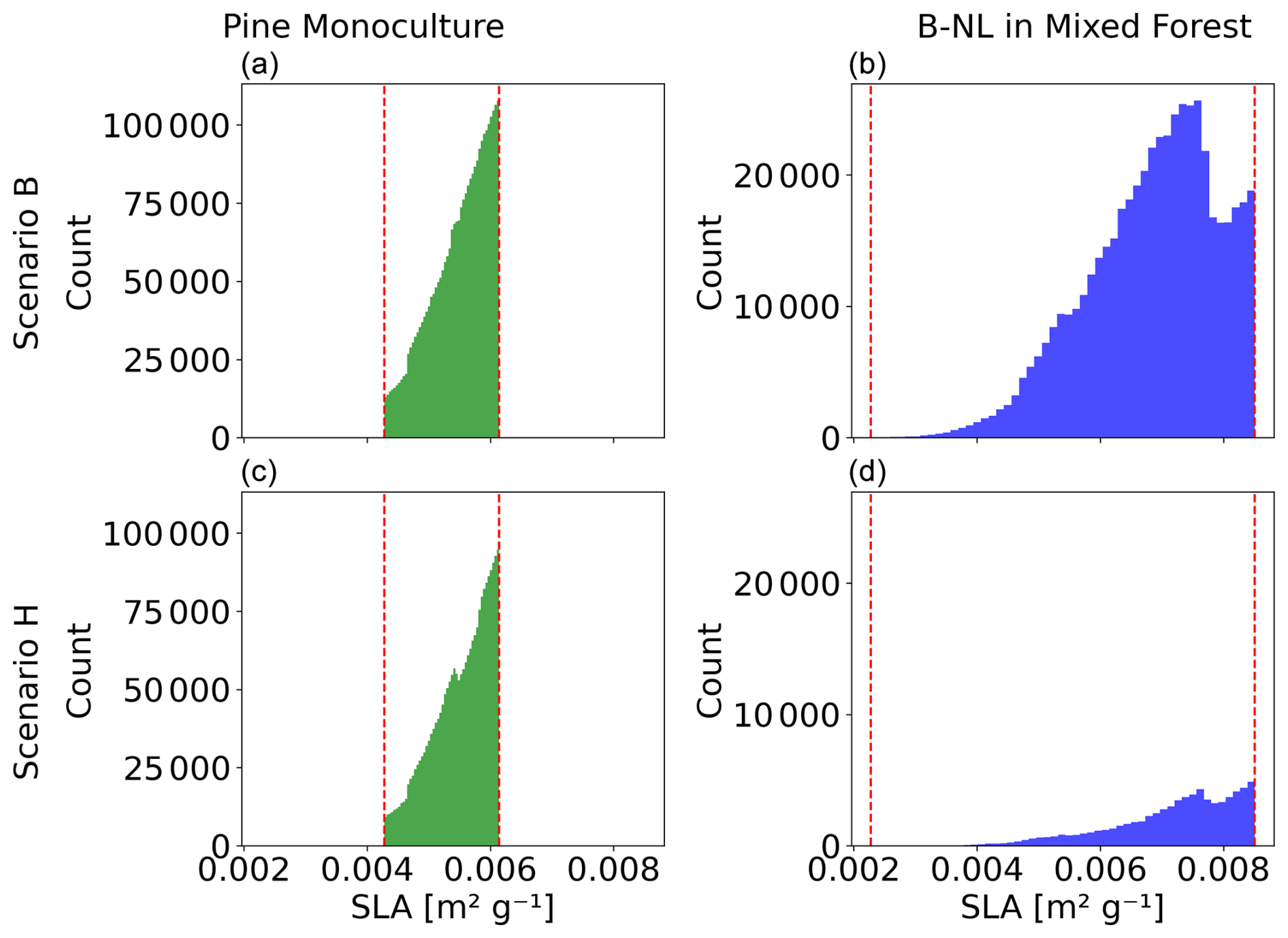
Figure C1Specific leaf area (SLA) distribution in the Berlin–Brandenburg study area for pine monoculture (a, c) and boreal needle-leaved (B-NL) trees growing in a mixed forest (b, d) in simulation year 800 of Scenario B (a, b) and Scenario C (c, d). Dashed red lines mark the limits of the SLA ranges set in the parametrization.
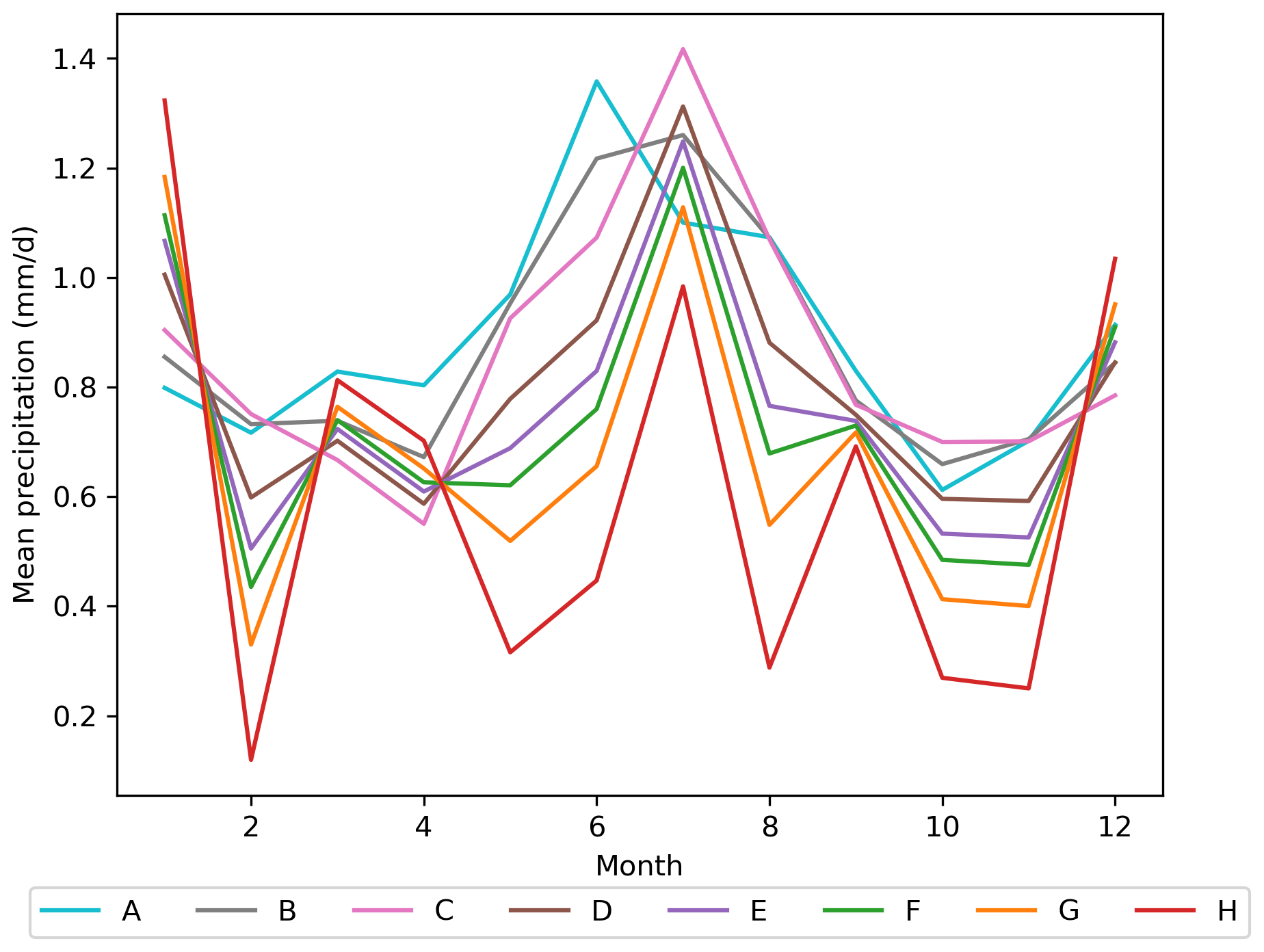
Figure D1Mean daily precipitation [mm d−1] for each month over all years in each Scenario A–H.
Programming code can be provided by the Potsdam Institute for Climate Impact Research (PIK e.V.) upon reasonable request and pending scientific review.
All data needed to evaluate the conclusions in the paper are present in the paper. The data can be provided by the Potsdam Institute for Climate Impact Research (PIK e.V.) upon reasonable request and pending scientific review.
JP, BT, KT, BS, and MB made the study design. KT, BS, SB, WvB, and MB developed the LPJml-FIT model version used in this study. JP carried out the simulations. JP, BT, KT, BS, MB, and SF analyzed and interpreted the simulation outputs. JP, BT, and KT prepared the manuscript with contributions from all co-authors.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
This article is part of the special issue “Current and future water-related risks in the Berlin–Brandenburg region”. It is not associated with a conference.
The authors thank Ainka Douglas for proofreading the English-language manuscript.
This research has been supported by the Einstein Stiftung Berlin (grant no ERU-2020-609), the Berlin University Alliance (grant no. ERU-2020-609), and the Fachagentur Nachwachsende Rohstoffe (FNR) (grant no. 2219WK39A4).
This paper was edited by Márk Somogyvári and reviewed by Miquel De Cáceres and one anonymous referee.
Aiba, M. and Nakashizuka, T.: Architectural differences associated with adult stature and wood density in 30 temperate tree species, Funct. Ecol., 23, 265–273, https://doi.org/10.1111/j.1365-2435.2008.01500.x, 2009.
Ali, A. and Yan, E.-R.: Relationships between biodiversity and carbon stocks in forest ecosystems: a systematic literature review, Trop. Ecol., 58, 1–14, 2017.
Ammer, C.: Unraveling the Importance of Inter- and Intraspecific Competition for the Adaptation of Forests to Climate Change, in: Progress in Botany, edited by: Cánovas, F. M., Lüttge, U., and Matyssek, R., Springer International Publishing, Cham, vol. 78, 345–367, https://doi.org/10.1007/124_2016_14, 2017.
Copernicus: Europe experiences widespread flooding and severe heatwaves in 2023, Copernicus, https://climate.copernicus.eu/europe-experiences-widespread-flooding-and-severe-heatwaves-2023 (last access: 12 August 2024), 2023.
Astigarraga, J., Esquivel-Muelbert, A., Ruiz-Benito, P., Rodríguez-Sánchez, F., Zavala, M. A., Vilà-Cabrera, A., Schelhaas, M.-J., Kunstler, G., Woodall, C. W., Cienciala, E., Dahlgren, J., Govaere, L., König, L. A., Lehtonen, A., Talarczyk, A., Liu, D., and Pugh, T. A. M.: Relative decline in density of Northern Hemisphere tree species in warm and arid regions of their climate niches, P. Natl. Acad. Sci. USA, 121, e2314899121, https://doi.org/10.1073/pnas.2314899121, 2024.
Austerlitz, F. and Garnier-Géré, P. H.: Modelling the impact of colonisation on genetic diversity and differentiation of forest trees: interaction of life cycle, pollen flow and seed long-distance dispersal, Heredity, 90, 282–290, https://doi.org/10.1038/sj.hdy.6800243, 2003.
Barredo, J. I., Brailescu, C., Teller, A., Sabatini, F. M., Mauri, A., and Janouskova, K.: Mapping and assessment of primary and old-growth forests in Europe, Publications Office of the European Union, https://doi.org/10.2760/797591, 2021.
Bart, F., Schmidt, B., Wang, X., Holtmann, A., Meier, F., Otto, M., and Scherer, D.: The Central Europe Refined analysis version 2 (CER v2): Evaluating three decades of high-resolution precipitation data for the Berlin-Brandenburg metropolitan region, Meteorol. Z., 339–363, https://doi.org/10.1127/metz/2024/1233, 2025.
Bauhus, J., Dieter, M., Farwig, N., Hafner, A., Kätzel, R., Kleinschmit, B., Lang, F., Lindner, M., Möhring, B., Müller, J., Niekisch, M., Richter, K., Schraml, U., and Seeling, U.: Die Anpassung von Wäldern und Waldwirtschaft an den Klimawandel: Gutachten des Wissenschaftlichen Beirates für Waldpolitik, Bundesministerium für Ernährung und Landwirtschaft, Berlin, ISBN 978-3-00-070408-6, 2021.
Bengtsson, J., Nilsson, S. G., Franc, A., and Menozzi, P.: Biodiversity, disturbances, ecosystem function and management of European forests, For. Ecol. Manag., 132, 39–50, https://doi.org/10.1016/S0378-1127(00)00378-9, 2000.
Bennett, A. C., McDowell, N. G., Allen, C. D., and Anderson-Teixeira, K. J.: Larger trees suffer most during drought in forests worldwide, Nat. Plants, 1, 1–5, https://doi.org/10.1038/nplants.2015.139, 2015.
Bigler, C. and Veblen, T. T.: Increased early growth rates decrease longevities of conifers in subalpine forests, Oikos, 118, 1130–1138, https://doi.org/10.1111/j.1600-0706.2009.17592.x, 2009.
Billing, M., Thonicke, K., Sakschewski, B., von Bloh, W., and Walz, A.: Future tree survival in European forests depends on understorey tree diversity, Sci. Rep., 12, 20750, https://doi.org/10.1038/s41598-022-25319-7, 2022.
Billing, M., Sakschewski, B., von Bloh, W., Vogel, J., and Thonicke, K.: “How to adapt forests?” – Exploring the role of leaf trait diversity for long-term forest biomass under new climate normals, Glob. Change Biol., 30, e17258, https://doi.org/10.1111/gcb.17258, 2024.
Black, B. A., Colbert, J. J., and Pederson, N.: Relationships between radial growth rates and lifespan within North American tree species, Écoscience, 15, 349–357, https://doi.org/10.2980/15-3-3149, 2008.
Bonebrake, T. C., Brown, C. J., Bell, J. D., Blanchard, J. L., Chauvenet, A., Champion, C., Chen, I.-C., Clark, T. D., Colwell, R. K., Danielsen, F., Dell, A. I., Donelson, J. M., Evengård, B., Ferrier, S., Frusher, S., Garcia, R. A., Griffis, R. B., Hobday, A. J., Jarzyna, M. A., Lee, E., Lenoir, J., Linnetved, H., Martin, V. Y., McCormack, P. C., McDonald, J., McDonald-Madden, E., Mitchell, N., Mustonen, T., Pandolfi, J. M., Pettorelli, N., Possingham, H., Pulsifer, P., Reynolds, M., Scheffers, B. R., Sorte, C. J. B., Strugnell, J. M., Tuanmu, M.-N., Twiname, S., Vergés, A., Villanueva, C., Wapstra, E., Wernberg, T., and Pecl, G. T.: Managing consequences of climate-driven species redistribution requires integration of ecology, conservation and social science, Biol. Rev., 93, 284–305, https://doi.org/10.1111/brv.12344, 2018.
Bouchard, E., Searle, E. B., Drapeau, P., Liang, J., Gamarra, J. G. P., Abegg, M., Alberti, G., Zambrano, A. A., Alvarez-Davila, E., Alves, L. F., Avitabile, V., Aymard, G., Bastin, J.-F., Birnbaum, P., Bongers, F., Bouriaud, O., Brancalion, P., Broadbent, E., Bussotti, F., Gatti, R. C., Češljar, G., Chisholm, C., Cienciala, E., Clark, C. J., Corral-Rivas, J. J., Crowther, T. W., Dayanandan, S., Decuyper, M., de Gasper, A. L., de-Miguel, S., Derroire, G., DeVries, B., Djordjević, I., Van Do, T., Dolezal, J., Fayle, T. M., Fridman, J., Frizzera, L., Gianelle, D., Hemp, A., Hérault, B., Herold, M., Imai, N., Jagodziński, A. M., Jaroszewicz, B., Jucker, T., Kepfer-Rojas, S., Keppel, G., Khan, M. L., Kim, H. S., Korjus, H., Kraxner, F., Laarmann, D., Lewis, S., Lu, H., Maitner, B. S., Marcon, E., Marshall, A. R., Mukul, S. A., Nabuurs, G.-J., Nava-Miranda, M. G., Parfenova, E. I., Park, M., Peri, P. L., Pfautsch, S., Phillips, O. L., Piedade, M. T. F., Piotto, D., Poulsen, J. R., Poulsen, A. D., Pretzsch, H., Reich, P. B., Rodeghiero, M., Rolim, S., Rovero, F., Saikia, P., Salas-Eljatib, C., Schall, P., Schepaschenko, D., Schöngart, J., Šebeň, V., Sist, P., Slik, F., Souza, A. F., Stereńczak, K., Svoboda, M., Tchebakova, N. M., ter Steege, H., Tikhonova, E. V., Usoltsev, V. A., Valladares, F., Viana, H., Vibrans, A. C., Wang, H.-F., Westerlund, B., Wiser, S. K., Wittmann, F., Wortel, V., Zawiła-Niedźwiecki, T., Zhou, M., Zhu, Z.-X., Zo-Bi, I. C., and Paquette, A.: Global patterns and environmental drivers of forest functional composition, Glob. Ecol. Biogeogr., 33, 303–324, https://doi.org/10.1111/geb.13790, 2024.
Brockerhoff, E. G., Barbaro, L., Castagneyrol, B., Forrester, D. I., Gardiner, B., González-Olabarria, J. R., Lyver, P. O., Meurisse, N., Oxbrough, A., Taki, H., Thompson, I. D., van der Plas, F., and Jactel, H.: Forest biodiversity, ecosystem functioning and the provision of ecosystem services, Biodivers. Conserv., 26, 3005–3035, https://doi.org/10.1007/s10531-017-1453-2, 2017.
Bundesministerium für Ernährung und Landwirtschaft: Ergebnisse der Waldzustandserhebung 2021, Bundesministerium für Ernährung und Landwirtschaft, 2021.
Büntgen, U., Urban, O., Krusic, P. J., Rybníček, M., Kolář, T., Kyncl, T., Ač, A., Koňasová, E., Čáslavský, J., Esper, J., Wagner, S., Saurer, M., Tegel, W., Dobrovolný, P., Cherubini, P., Reinig, F., and Trnka, M.: Recent European drought extremes beyond Common Era background variability, Nat. Geosci., 14, 190–196, https://doi.org/10.1038/s41561-021-00698-0, 2021.
Buras, A. and Menzel, A.: Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios, Front. Plant Sci., 9, 88–100, https://doi.org/10.3389/fpls.2018.01986, 2019.
Buras, A., Rammig, A., and Zang, C. S.: Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003, Biogeosciences, 17, 1655–1672, https://doi.org/10.5194/bg-17-1655-2020, 2020.
Chakraborty, D., Ciceu, A., Ballian, D., Benito Garzón, M., Bolte, A., Bozic, G., Buchacher, R., Čepl, J., Cremer, E., Ducousso, A., Gaviria, J., George, J. P., Hardtke, A., Ivankovic, M., Klisz, M., Kowalczyk, J., Kremer, A., Lstibůrek, M., Longauer, R., Mihai, G., Nagy, L., Petkova, K., Popov, E., Schirmer, R., Skrøppa, T., Solvin, T. M., Steffenrem, A., Stejskal, J., Stojnic, S., Volmer, K., and Schueler, S.: Assisted tree migration can preserve the European forest carbon sink under climate change, Nat. Clim. Change, 14, 845–852, https://doi.org/10.1038/s41558-024-02080-5, 2024.
Charru, M., Seynave, I., Hervé, J.-C., Bertrand, R., and Bontemps, J.-D.: Recent growth changes in Western European forests are driven by climate warming and structured across tree species climatic habitats, Ann. For. Sci., 74, 1–34, https://doi.org/10.1007/s13595-017-0626-1, 2017.
Chave, J., Coomes, D., Jansen, S., Lewis, S. L., Swenson, N. G., and Zanne, A. E.: Towards a worldwide wood economics spectrum, Ecol. Lett., 12, 351–366, https://doi.org/10.1111/j.1461-0248.2009.01285.x, 2009.
Chen, I.-C., Hill, J. K., Ohlemüller, R., Roy, D. B., and Thomas, C. D.: Rapid Range Shifts of Species Associated with High Levels of Climate Warming, Science, 333, 1024–1026, https://doi.org/10.1126/science.1206432, 2011.
Cheng, J., Zhang, M., Yan, X., Zhang, C., Zhang, J., and Luo, Y.: Effects of Seed Size and Frequency on Seed Dispersal and Predation by Small Mammals, Biology, 13, 353, https://doi.org/10.3390/biology13050353, 2024.
Chisholm, R. A. and Dutta Gupta, T.: A critical assessment of the biodiversity–productivity relationship in forests and implications for conservation, Oecologia, 201, 887–900, https://doi.org/10.1007/s00442-023-05363-4, 2023.
Decarsin, R., Guillemot, J., le Maire, G., Blondeel, H., Meredieu, C., Achard, E., Bonal, D., Cochard, H., Corso, D., Delzon, S., Doucet, Z., Druel, A., Grossiord, C., Torres-Ruiz, J. M., Bauhus, J., Godbold, D. L., Hajek, P., Jactel, H., Jensen, J., Mereu, S., Ponette, Q., Rewald, B., Ruffault, J., Sandén, H., Scherer-Lorenzen, M., Serrano-León, H., Simioni, G., Verheyen, K., Werner, R., and Martin-StPaul, N.: Tree drought–mortality risk depends more on intrinsic species resistance than on stand species diversity, Glob. Change Biol., 30, e17503, https://doi.org/10.1111/gcb.17503, 2024.
Dittmann, D., Seelig, A. H., Thalmann, M., Wilkes, T., Junghans, V., Zahn, D., Klitzke, S., Peters, A., Haberkamp, J., Reemtsma, T., and Ruhl, A. S.: Potential and risks of water reuse in Brandenburg (Germany) – an interdisciplinary case study, Water Reuse, 14, 1–15, https://doi.org/10.2166/wrd.2024.081, 2024.
FAO and IIASA: Harmonized World Soil Database version 2.0, FAO, International Institute for Applied Systems Analysis (IIASA), ISBN 978-92-5-137499-3, 2023.
Fei, S., Desprez, J. M., Potter, K. M., Jo, I., Knott, J. A., and Oswalt, C. M.: Divergence of species responses to climate change, Sci. Adv., 3, e1603055, https://doi.org/10.1126/sciadv.1603055, 2017.
Fei, S., Jo, I., Guo, Q., Wardle, D. A., Fang, J., Chen, A., Oswalt, C. M., and Brockerhoff, E. G.: Impacts of climate on the biodiversity-productivity relationship in natural forests, Nat. Commun., 9, 5436, https://doi.org/10.1038/s41467-018-07880-w, 2018.
Forrester, D. I., Bonal, D., Dawud, S., Gessler, A., Granier, A., Pollastrini, M., and Grossiord, C.: Drought responses by individual tree species are not often correlated with tree species diversity in European forests, J. Appl. Ecol., 53, 1725–1734, https://doi.org/10.1111/1365-2664.12745, 2016.
Förster, A., Culmsee, H., and Leuschner, C.: Thinned northern German Scots pine forests have a low carbon storage and uptake potential in comparison to naturally developing beech forests, For. Ecol. Manag., 479, 118575, https://doi.org/10.1016/j.foreco.2020.118575, 2021.
Galván, J. D., Camarero, J. J., and Gutiérrez, E.: Seeing the trees for the forest: drivers of individual growth responses to climate in pinus uncinata mountain forests, J. Ecol., 102, 1244–1257, https://doi.org/10.1111/1365-2745.12268, 2014.
Gerber, S., Chadœuf, J., Gugerli, F., Lascoux, M., Buiteveld, J., Cottrell, J., Dounavi, A., Fineschi, S., Forrest, L. L., Fogelqvist, J., Goicoechea, P. G., Jensen, J. S., Salvini, D., Vendramin, G. G., and Kremer, A.: High Rates of Gene Flow by Pollen and Seed in Oak Populations across Europe, PLOS ONE, 9, e85130, https://doi.org/10.1371/journal.pone.0085130, 2014.
Granda, E., Gazol, A., and Camarero, J. J.: Functional diversity differently shapes growth resilience to drought for co-existing pine species, J. Veg. Sci., 29, 265–275, https://doi.org/10.1111/jvs.12617, 2018.
Greenwood, S., Ruiz-Benito, P., Martínez-Vilalta, J., Lloret, F., Kitzberger, T., Allen, C. D., Fensham, R., Laughlin, D. C., Kattge, J., Bönisch, G., Kraft, N. J. B., and Jump, A. S.: Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area, Ecol. Lett., 20, 539–553, https://doi.org/10.1111/ele.12748, 2017.
Gregor, K., Knoke, T., Krause, A., Reyer, C. P. O., Lindeskog, M., Papastefanou, P., Smith, B., Lansø, A.-S., and Rammig, A.: Trade-Offs for Climate-Smart Forestry in Europe Under Uncertain Future Climate, Earths Future, 10, e2022EF002796, https://doi.org/10.1029/2022EF002796, 2022.
Gutsch, M., Lasch-Born, P., Kollas, C., Suckow, F., and Reyer, C. P. O.: Balancing trade-offs between ecosystem services in Germany's forests under climate change, Environ. Res. Lett., 13, 045012, https://doi.org/10.1088/1748-9326/aab4e5, 2018.
Haberstroh, S., Werner, C., Grün, M., Kreuzwieser, J., Seifert, T., Schindler, D., and Christen, A.: Central European 2018 hot drought shifts scots pine forest to its tipping point, Plant Biol., 24, 1186–1197, https://doi.org/10.1111/plb.13455, 2022.
Hanewinkel, M., Lessa Derci Augustynczik, A., and Yousefpour, R.: Climate-Smart Forestry Case Study: Germany, in: Forest Bioeconomy and Climate Change, edited by: Hetemäki, L., Kangas, J., and Peltola, H., Springer International Publishing, Cham, 197–209, https://doi.org/10.1007/978-3-030-99206-4_12, 2022.
Hannerz, M., Ekberg, I., and Norell, L.: Variation in Chilling Requirements for Completing Bud Rest Between Provenances of Norway Spruce, Silvae Genetica 52, 161–168, 2003.
Hisano, M., Chen, H. Y. H., Searle, E. B., and Reich, P. B.: Species-rich boreal forests grew more and suffered less mortality than species-poor forests under the environmental change of the past half-century, Ecol. Lett., 22, 999–1008, https://doi.org/10.1111/ele.13259, 2019.
Houšková, K., Klepárník, J., and Mauer, O.: How to accelerate the germination of Scots pine and Norway spruce seeds?, J. For. Sci., 67, 134–142, https://doi.org/10.17221/133/2020-JFS, 2021.
Húdoková, H., Petrik, P., Petek-Petrik, A., Konôpková, A., Leštianska, A., Střelcová, K., Kmeť, J., and Kurjak, D.: Heat-stress response of photosystem II in five ecologically important tree species of European temperate forests, Biologia (Bratisl.), 77, 671–680, https://doi.org/10.1007/s11756-021-00958-9, 2022.
Johnson, S. E. and Abrams, M. D.: Age class, longevity and growth rate relationships: protracted growth increases in old trees in the eastern United States, Tree Physiol., 29, 1317–1328, https://doi.org/10.1093/treephys/tpp068, 2009.
Kallarackal, J. and Ramírez, F.: Functional Correlations of Wood Density, in: Wood Density: Functional Trait in Plants, edited by: Kallarackal, J. and Ramírez, F., Springer International Publishing, Cham, 21–40, https://doi.org/10.1007/978-3-031-61030-1_3, 2024.
Kattge, J., Díaz, S., Lavorel, S., Prentice, I. C., Leadley, P., Bönisch, G., Garnier, E., Westoby, M., Reich, P. B., Wright, I. J., Cornelissen, J. H. C., Violle, C., Harrison, S. P., Van BODEGOM, P. M., Reichstein, M., Enquist, B. J., Soudzilovskaia, N. A., Ackerly, D. D., Anand, M., Atkin, O., Bahn, M., Baker, T. R., Baldocchi, D., Bekker, R., Blanco, C. C., Blonder, B., Bond, W. J., Bradstock, R., Bunker, D. E., Casanoves, F., Cavender-Bares, J., Chambers, J. Q., Chapin Iii, F. S., Chave, J., Coomes, D., Cornwell, W. K., Craine, J. M., Dobrin, B. H., Duarte, L., Durka, W., Elser, J., Esser, G., Estiarte, M., Fagan, W. F., Fang, J., Fernández-Méndez, F., Fidelis, A., Finegan, B., Flores, O., Ford, H., Frank, D., Freschet, G. T., Fyllas, N. M., Gallagher, R. V., Green, W. A., Gutierrez, A. G., Hickler, T., Higgins, S. I., Hodgson, J. G., Jalili, A., Jansen, S., Joly, C. A., Kerkhoff, A. J., Kirkup, D., Kitajima, K., Kleyer, M., Klotz, S., Knops, J. M. H., Kramer, K., Kühn, I., Kurokawa, H., Laughlin, D., Lee, T. D., Leishman, M., Lens, F., Lenz, T., Lewis, S. L., Lloyd, J., Llusià, J., Louault, F., Ma, S., Mahecha, M. D., Manning, P., Massad, T., Medlyn, B. E., Messier, J., Moles, A. T., Müller, S. C., Nadrowski, K., Naeem, S., Niinemets, Ü., Nöllert, S., Nüske, A., Ogaya, R., Oleksyn, J., Onipchenko, V. G., Onoda, Y., Ordoñez, J., Overbeck, G., Ozinga, W. A., Patiño, S., Paula, S., Pausas, J. G., Peñuelas, J., Phillips, O. L., Pillar, V., Poorter, H., Poorter, L., Poschlod, P., Prinzing, A., Proulx, R., Rammig, A., Reinsch, S., Reu, B., Sack, L., Salgado-Negret, B., Sardans, J., Shiodera, S., Shipley, B., Siefert, A., Sosinski, E., Soussana, J.-F., Swaine, E., Swenson, N., Thompson, K., Thornton, P., Waldram, M., Weiher, E., White, M., White, S., Wright, S. J., Yguel, B., Zaehle, S., Zanne, A. E., Wirth, C.: TRY – a global database of plant traits, Glob. Change Biol., 17, 2905–2935, https://doi.org/10.1111/j.1365-2486.2011.02451.x, 2011.
Kim, M., Lee, S., Lee, S., Yi, K., Kim, H.-S., Chung, S., Chung, J., Kim, H. S., and Yoon, T. K.: Seed Dispersal Models for Natural Regeneration: A Review and Prospects, Forests, 13, 659, https://doi.org/10.3390/f13050659, 2022.
Lamentowicz, M., Marcisz, K., Guzowski, P., Gałka, M., Diaconu, A.-C., and Kołaczek, P.: How Joannites' economy eradicated primeval forest and created anthroecosystems in medieval Central Europe, Sci. Rep., 10, 18775, https://doi.org/10.1038/s41598-020-75692-4, 2020.
Land Brandenburg: Waldprogramm 2011, Land Brandenburg, 2011.
Land Brandenburg: WALDZUSTANDSBERICHT 2023 DES LANDES BRANDENBURG, Land Brandenburg, 2023.
Landeskompetenzentrum Forst Eberswalde: Waldbrandstatistik 2018, Landeskompetenzentrum Forst Eberswalde, 2018.
Laurance, W. F., Nascimento, H. E. M., Laurance, S. G., Condit, R., D'Angelo, S., and Andrade, A.: Inferred longevity of Amazonian rainforest trees based on a long-term demographic study, For. Ecol. Manag., 190, 131–143, https://doi.org/10.1016/j.foreco.2003.09.011, 2004.
Lehsten, V., Mischurow, M., Lindström, E., Lehsten, D., and Lischke, H.: LPJ-GM 1.0: simulating migration efficiently in a dynamic vegetation model, Geosci. Model Dev., 12, 893–908, https://doi.org/10.5194/gmd-12-893-2019, 2019.
Lenoir, J. and Svenning, J.-C.: Climate-related range shifts – a global multidimensional synthesis and new research directions, Ecography, 38, 15–28, https://doi.org/10.1111/ecog.00967, 2015.
LfU Brandenburg: Klimawandelmonitoring im Land Brandenburg – Aktualisierungsbericht, LfU Brandenburg, 2021.
LfU Brandenburg: Klimawandel in Brandenburg, LfU Brandenburg, 2022.
Martinez del Castillo, E., Zang, C. S., Buras, A., Hacket-Pain, A., Esper, J., Serrano-Notivoli, R., Hartl, C., Weigel, R., Klesse, S., Resco de Dios, V., Scharnweber, T., Dorado-Liñán, I., van der Maaten-Theunissen, M., van der Maaten, E., Jump, A., Mikac, S., Banzragch, B.-E., Beck, W., Cavin, L., Claessens, H., Čada, V., Čufar, K., Dulamsuren, C., Gričar, J., Gil-Pelegrín, E., Janda, P., Kazimirovic, M., Kreyling, J., Latte, N., Leuschner, C., Longares, L. A., Menzel, A., Merela, M., Motta, R., Muffler, L., Nola, P., Petritan, A. M., Petritan, I. C., Prislan, P., Rubio-Cuadrado, Á., Rydval, M., Stajić, B., Svoboda, M., Toromani, E., Trotsiuk, V., Wilmking, M., Zlatanov, T., and de Luis, M.: Climate-change-driven growth decline of European beech forests, Commun. Biol., 5, 1–9, https://doi.org/10.1038/s42003-022-03107-3, 2022.
Milad, M., Schaich, H., and Konold, W.: How is adaptation to climate change reflected in current practice of forest management and conservation? A case study from Germany, Biodivers. Conserv., 22, 1181–1202, https://doi.org/10.1007/s10531-012-0337-8, 2013.
Mori, A. S., Furukawa, T., and Sasaki, T.: Response diversity determines the resilience of ecosystems to environmental change, Biol. Rev., 88, 349–364, https://doi.org/10.1111/brv.12004, 2013.
Morin, X., Fahse, L., Scherer-Lorenzen, M., and Bugmann, H.: Tree species richness promotes productivity in temperate forests through strong complementarity between species, Ecol. Lett., 14, 1211–1219, https://doi.org/10.1111/j.1461-0248.2011.01691.x, 2011.
Nabais, C., Hansen, J. K., David-Schwartz, R., Klisz, M., López, R., and Rozenberg, P.: The effect of climate on wood density: What provenance trials tell us?, For. Ecol. Manag., 408, 148–156, https://doi.org/10.1016/j.foreco.2017.10.040, 2018.
Nelson, R. A., Francis, E. J., Berry, J. A., Cornwell, W. K., and Anderegg, L. D. L.: The Role of Climate Niche, Geofloristic History, Habitat Preference, and Allometry on Wood Density within a California Plant Community, Forests, 11, 105, https://doi.org/10.3390/f11010105, 2020.
Niedertscheider, M., Kuemmerle, T., Müller, D., and Erb, K.-H.: Exploring the effects of drastic institutional and socio-economic changes on land system dynamics in Germany between 1883 and 2007, Glob. Environ. Change Hum. Policy Dimens., 28, 98–108, https://doi.org/10.1016/j.gloenvcha.2014.06.006, 2014.
Ostberg, S., Müller, C., Heinke, J., and Schaphoff, S.: LandInG 1.0: a toolbox to derive input datasets for terrestrial ecosystem modelling at variable resolutions from heterogeneous sources, Geosci. Model Dev., 16, 3375–3406, https://doi.org/10.5194/gmd-16-3375-2023, 2023.
Pardos, M., del Río, M., Pretzsch, H., Jactel, H., Bielak, K., Bravo, F., Brazaitis, G., Defossez, E., Engel, M., Godvod, K., Jacobs, K., Jansone, L., Jansons, A., Morin, X., Nothdurft, A., Oreti, L., Ponette, Q., Pach, M., Riofrío, J., Ruíz-Peinado, R., Tomao, A., Uhl, E., and Calama, R.: The greater resilience of mixed forests to drought mainly depends on their composition: Analysis along a climate gradient across Europe, For. Ecol. Manag., 481, 118687, https://doi.org/10.1016/j.foreco.2020.118687, 2021.
Parmesan, C. and Yohe, G.: A globally coherent fingerprint of climate change impacts across natural systems, Nature, 421, 37–42, https://doi.org/10.1038/nature01286, 2003.
Pelletier, J. D., Broxton, P. D., Hazenberg, P., Zeng, X., Troch, P. A., Niu, G.-Y., Williams, Z., Brunke, M. A., and Gochis, D.: A gridded global data set of soil, intact regolith, and sedimentary deposit thicknesses for regional and global land surface modeling, J. Adv. Model. Earth Syst., 8, 41–65, https://doi.org/10.1002/2015MS000526, 2016.
Piovesan, G., Biondi, F., Filippo, A. D., Alessandrini, A., and Maugeri, M.: Drought-driven growth reduction in old beech (Fagus sylvatica L.) forests of the central Apennines, Italy, Glob. Change Biol., 14, 1265–1281, https://doi.org/10.1111/j.1365-2486.2008.01570.x, 2008.
Poorter, L., McDonald, I., Alarcón, A., Fichtler, E., Licona, J.-C., Peña-Claros, M., Sterck, F., Villegas, Z., and Sass-Klaassen, U.: The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species, New Phytol., 185, 481–492, https://doi.org/10.1111/j.1469-8137.2009.03092.x, 2010.
Pradhan, P., Seydewitz, T., Zhou, B., Lüdeke, M. K. B., and Kropp, J. P.: Climate Extremes are Becoming More Frequent, Co-occurring, and Persistent in Europe, Anthr. Sci., 1, 264–277, https://doi.org/10.1007/s44177-022-00022-4, 2022.
Pretzsch, H. and Biber, P.: Tree species mixing can increase maximum stand density, Can. J. For. Res., 46, 1179–1193, https://doi.org/10.1139/cjfr-2015-0413, 2016.
Pretzsch, H. and Forrester, D. I.: Stand Dynamics of Mixed-Species Stands Compared with Monocultures, in: Mixed-Species Forests: Ecology and Management, edited by: Pretzsch, H., Forrester, D. I., and Bauhus, J., Springer, Berlin, Heidelberg, 117–209, https://doi.org/10.1007/978-3-662-54553-9_4, 2017.
Pretzsch, H. and Schütze, G.: Effect of tree species mixing on the size structure, density, and yield of forest stands, Eur. J. For. Res., 135, 1–22, https://doi.org/10.1007/s10342-015-0913-z, 2016.
Pretzsch, H., Schütze, G., and Uhl, E.: Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation, Plant Biol., 15, 483–495, https://doi.org/10.1111/j.1438-8677.2012.00670.x, 2013.
Pretzsch, H., Biber, P., Schütze, G., Uhl, E., and Rötzer, T.: Forest stand growth dynamics in Central Europe have accelerated since 1870, Nat. Commun., 5, 4967, https://doi.org/10.1038/ncomms5967, 2014.
Pretzsch, H., Hilmers, T., Biber, P., Avdagić, A., Binder, F., Bončina, A., Bosela, M., Dobor, L., Forrester, D. I., Lévesque, M., Ibrahimspahić, A., Nagel, T. A., del Río, M., Sitkova, Z., Schütze, G., Stajić, B., Stojanović, D., Uhl, E., Zlatanov, T., and Tognetti, R.: Evidence of elevation-specific growth changes of spruce, fir, and beech in European mixed mountain forests during the last three centuries, Can. J. For. Res., 50, 689–703, https://doi.org/10.1139/cjfr-2019-0368, 2020.
Pretzsch, H., del Río, M., Arcangeli, C., Bielak, K., Dudzinska, M., Forrester, D. I., Klädtke, J., Kohnle, U., Ledermann, T., Matthews, R., Nagel, J., Nagel, R., Ningre, F., Nord-Larsen, T., and Biber, P.: Forest growth in Europe shows diverging large regional trends, Sci. Rep., 13, 15373, https://doi.org/10.1038/s41598-023-41077-6, 2023.
Rabasa, S. G., Granda, E., Benavides, R., Kunstler, G., Espelta, J. M., Ogaya, R., Peñuelas, J., Scherer-Lorenzen, M., Gil, W., Grodzki, W., Ambrozy, S., Bergh, J., Hódar, J. A., Zamora, R., and Valladares, F.: Disparity in elevational shifts of European trees in response to recent climate warming, Glob. Change Biol., 19, 2490–2499, https://doi.org/10.1111/gcb.12220, 2013.
Rampart, M.: Thermal time models and predictions for germination of six provenances of Scots pine (Pinus sylvestris) in southern England, University of Tlemcen – ALGERIA: Agriculture and Forestry Journal, 2, 81–88, E-ISSN 2602-5795, 2018.
Reinermann, S., Gessner, U., Asam, S., Kuenzer, C., and Dech, S.: The Effect of Droughts on Vegetation Condition in Germany: An Analysis Based on Two Decades of Satellite Earth Observation Time Series and Crop Yield Statistics, Remote Sens., 11, 1783, https://doi.org/10.3390/rs11151783, 2019.
Richards, A. E., Forrester, D. I., Bauhus, J., and Scherer-Lorenzen, M.: The influence of mixed tree plantations on the nutrition of individual species: a review, Tree Physiol., 30, 1192–1208, https://doi.org/10.1093/treephys/tpq035, 2010.
Rubenstein, M. A., Weiskopf, S. R., Carter, S. L., Eaton, M. J., Johnson, C., Lynch, A. J., Miller, B. W., Morelli, T. L., Rodriguez, M. A., Terando, A., and Thompson, L. M.: Do empirical observations support commonly-held climate change range shift hypotheses? A systematic review protocol, Environ. Evid., 9, 10, https://doi.org/10.1186/s13750-020-00194-9, 2020.
Rubenstein, M. A., Weiskopf, S. R., Bertrand, R., Carter, S. L., Comte, L., Eaton, M. J., Johnson, C. G., Lenoir, J., Lynch, A. J., Miller, B. W., Morelli, T. L., Rodriguez, M. A., Terando, A., and Thompson, L. M.: Climate change and the global redistribution of biodiversity: substantial variation in empirical support for expected range shifts, Environ. Evid., 12, 7, https://doi.org/10.1186/s13750-023-00296-0, 2023.
Ryan, M. G. and Yoder, B. J.: Hydraulic Limits to Tree Height and Tree Growth, BioScience, 47, 235–242, https://doi.org/10.2307/1313077, 1997.
Sakschewski, B., von Bloh, W., Boit, A., Rammig, A., Kattge, J., Poorter, L., Peñuelas, J., and Thonicke, K.: Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model, Glob. Change Biol., 21, 2711–2725, https://doi.org/10.1111/gcb.12870, 2015.
Sakschewski, B., von Bloh, W., Drüke, M., Sörensson, A. A., Ruscica, R., Langerwisch, F., Billing, M., Bereswill, S., Hirota, M., Oliveira, R. S., Heinke, J., and Thonicke, K.: Variable tree rooting strategies are key for modelling the distribution, productivity and evapotranspiration of tropical evergreen forests, Biogeosciences, 18, 4091–4116, https://doi.org/10.5194/bg-18-4091-2021, 2021.
Schaphoff, S., Forkel, M., Müller, C., Knauer, J., von Bloh, W., Gerten, D., Jägermeyr, J., Lucht, W., Rammig, A., Thonicke, K., and Waha, K.: LPJmL4 – a dynamic global vegetation model with managed land – Part 2: Model evaluation, Geosci. Model Dev., 11, 1377–1403, https://doi.org/10.5194/gmd-11-1377-2018, 2018.
Schmied, G., Hilmers, T., Mellert, K.-H., Uhl, E., Buness, V., Ambs, D., Steckel, M., Biber, P., Šeho, M., Hoffmann, Y.-D., and Pretzsch, H.: Nutrient regime modulates drought response patterns of three temperate tree species, Sci. Total Environ., 868, 161601, https://doi.org/10.1016/j.scitotenv.2023.161601, 2023.
Schuldt, B., Buras, A., Arend, M., Vitasse, Y., Beierkuhnlein, C., Damm, A., Gharun, M., Grams, T. E. E., Hauck, M., Hajek, P., Hartmann, H., Hiltbrunner, E., Hoch, G., Holloway-Phillips, M., Körner, C., Larysch, E., Lübbe, T., Nelson, D. B., Rammig, A., Rigling, A., Rose, L., Ruehr, N. K., Schumann, K., Weiser, F., Werner, C., Wohlgemuth, T., Zang, C. S., and Kahmen, A.: A first assessment of the impact of the extreme 2018 summer drought on Central European forests, Basic Appl. Ecol., 45, 86–103, https://doi.org/10.1016/j.baae.2020.04.003, 2020.
Serra-Maluquer, X., Gazol, A., Anderegg, W. R. L., Martínez-Vilalta, J., Mencuccini, M., and Camarero, J. J.: Wood density and hydraulic traits influence species' growth response to drought across biomes, Glob. Change Biol., 28, 3871–3882, https://doi.org/10.1111/gcb.16123, 2022.
Spathelf, P. and Ammer, C.: Forest management of scots pine (Pinus sylvestris L) in northern Germany-a brief review of the history and current trends, Forstarchiv, 86, 59–66, https://doi.org/10.4432/0300-4112-86-59, 2015.
Sterk, M., Gort, G., Klimkowska, A., van Ruijven, J., van Teeffelen, A. J. A., and Wamelink, G. W. W.: Assess ecosystem resilience: Linking response and effect traits to environmental variability, Ecol. Indic., 30, 21–27, https://doi.org/10.1016/j.ecolind.2013.02.001, 2013.
Suding, K. N., Lavorel, S., Chapin Iii, F. S., Cornelissen, J. H. C., Díaz, S., Garnier, E., Goldberg, D., Hooper, D. U., Jackson, S. T., and Navas, M.-L.: Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants, Glob. Change Biol., 14, 1125–1140, https://doi.org/10.1111/j.1365-2486.2008.01557.x, 2008.
Swenson, N. G. and Enquist, B. J.: Ecological and evolutionary determinants of a key plant functional trait: wood density and its community-wide variation across latitude and elevation, Am. J. Bot., 94, 451–459, https://doi.org/10.3732/ajb.94.3.451, 2007.
Thompson, P. L. and Fronhofer, E. A.: The conflict between adaptation and dispersal for maintaining biodiversity in changing environments, P. Natl. Acad. Sci. USA, 116, 21061–21067, https://doi.org/10.1073/pnas.1911796116, 2019.
Thonicke, K., Billing, M., von Bloh, W., Sakschewski, B., Niinemets, Ü., Peñuelas, J., Cornelissen, J. H. C., Onoda, Y., van Bodegom, P., Schaepman, M. E., Schneider, F. D., and Walz, A.: Simulating functional diversity of European natural forests along climatic gradients, J. Biogeogr., 47, 1069–1085, https://doi.org/10.1111/jbi.13809, 2020.
Tillman-Sutela, E. and Kauppi, A.: The morphological background to imbibition in seeds ofPinus sylvestris L. of different provenances, Trees, 9, 123–133, https://doi.org/10.1007/BF02418201, 1995.
Treydte, K., Liu, L., Padrón, R. S., Martínez-Sancho, E., Babst, F., Frank, D. C., Gessler, A., Kahmen, A., Poulter, B., Seneviratne, S. I., Stegehuis, A. I., Wilson, R., Andreu-Hayles, L., Bale, R., Bednarz, Z., Boettger, T., Berninger, F., Büntgen, U., Daux, V., Dorado-Liñán, I., Esper, J., Friedrich, M., Gagen, M., Grabner, M., Grudd, H., Gunnarsson, B. E., Gutiérrez, E., Hafner, P., Haupt, M., Hilasvuori, E., Heinrich, I., Helle, G., Jalkanen, R., Jungner, H., Kalela-Brundin, M., Kessler, A., Kirchhefer, A., Klesse, S., Krapiec, M., Levanič, T., Leuenberger, M., Linderholm, H. W., McCarroll, D., Masson-Delmotte, V., Pawelczyk, S., Pazdur, A., Planells, O., Pukiene, R., Rinne-Garmston, K. T., Robertson, I., Saracino, A., Saurer, M., Schleser, G. H., Seftigen, K., Siegwolf, R. T. W., Sonninen, E., Stievenard, M., Szychowska-Krapiec, E., Szymaszek, M., Todaro, L., Waterhouse, J. S., Weigl-Kuska, M., Weigt, R. B., Wimmer, R., Woodley, E. J., Vitas, A., Young, G., and Loader, N. J.: Recent human-induced atmospheric drying across Europe unprecedented in the last 400 years, Nat. Geosci., 17, 58–65, https://doi.org/10.1038/s41561-023-01335-8, 2024.
Valladares, F. and Sánchez-Gómez, D.: Ecophysiological Traits Associated with Drought in Mediterranean Tree Seedlings: Individual Responses versus Interspecific Trends in Eleven Species, Plant Biol., 8, 688–697, https://doi.org/10.1055/s-2006-924107, 2006.
van der Wiel, K., Lenderink, G., and de Vries, H.: Physical storylines of future European drought events like 2018 based on ensemble climate modelling, Weather Clim. Extrem., 33, 100350, https://doi.org/10.1016/j.wace.2021.100350, 2021.
Watson, J. E. M., Evans, T., Venter, O., Williams, B., Tulloch, A., Stewart, C., Thompson, I., Ray, J. C., Murray, K., Salazar, A., McAlpine, C., Potapov, P., Walston, J., Robinson, J. G., Painter, M., Wilkie, D., Filardi, C., Laurance, W. F., Houghton, R. A., Maxwell, S., Grantham, H., Samper, C., Wang, S., Laestadius, L., Runting, R. K., Silva-Chávez, G. A., Ervin, J., and Lindenmayer, D.: The exceptional value of intact forest ecosystems, Nat. Ecol. Evol., 2, 599–610, https://doi.org/10.1038/s41559-018-0490-x, 2018.
Weigel, R., Bat-Enerel, B., Dulamsuren, C., Muffler, L., Weithmann, G., and Leuschner, C.: Summer drought exposure, stand structure, and soil properties jointly control the growth of European beech along a steep precipitation gradient in northern Germany, Glob. Change Biol., 29, 763–779, https://doi.org/10.1111/gcb.16506, 2023.
Wessely, J., Essl, F., Fiedler, K., Gattringer, A., Hülber, B., Ignateva, O., Moser, D., Rammer, W., Dullinger, S., and Seidl, R.: A climate-induced tree species bottleneck for forest management in Europe, Nat. Ecol. Evol., 8, 1109–1117, https://doi.org/10.1038/s41559-024-02406-8, 2024.
Williams, A. P., Allen, C. D., Macalady, A. K., Griffin, D., Woodhouse, C. A., Meko, D. M., Swetnam, T. W., Rauscher, S. A., Seager, R., Grissino-Mayer, H. D., Dean, J. S., Cook, E. R., Gangodagamage, C., Cai, M., and McDowell, N. G.: Temperature as a potent driver of regional forest drought stress and tree mortality, Nat. Clim. Change, 3, 292–297, https://doi.org/10.1038/nclimate1693, 2013.
Yachi, S. and Loreau, M.: Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis, P. Natl. Acad. Sci. USA, 96, 1463–1468, https://doi.org/10.1073/pnas.96.4.1463, 1999.
Zani, D., Lischke, H., and Lehsten, V.: Climate and dispersal limitation drive tree species range shifts in post-glacial Europe: results from dynamic simulations, Front. Ecol. Evol., 11, 1321104, https://doi.org/10.3389/fevo.2023.1321104, 2023.
Zhang, S.-B., Slik, J. W. F., Zhang, J.-L., and Cao, K.-F.: Spatial patterns of wood traits in China are controlled by phylogeny and the environment, Glob. Ecol. Biogeogr., 20, 241–250, https://doi.org/10.1111/j.1466-8238.2010.00582.x, 2011.
Zhang, Y. and Chen, H. Y. H.: Individual size inequality links forest diversity and above-ground biomass, J. Ecol., 103, 1245–1252, https://doi.org/10.1111/1365-2745.12425, 2015.
Zhu, K., Woodall, C. W., Ghosh, S., Gelfand, A. E., and Clark, J. S.: Dual impacts of climate change: forest migration and turnover through life history, Glob. Change Biol., 20, 251–264, https://doi.org/10.1111/gcb.12382, 2014.
Zscheischler, J. and Fischer, E. M.: The record-breaking compound hot and dry 2018 growing season in Germany, Weather Clim. Extrem., 29, 100270, https://doi.org/10.1016/j.wace.2020.100270, 2020.
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusions
- Appendix A: Selected characteristics of plant functional types and Pinus sylvestris parametrization
- Appendix B: Biomass, temperature, and maximum climatic water deficit maps for Berlin–Brandenburg
- Appendix C: Examples of SLA distributions
- Appendix D: Precipitation seasonality
- Code availability
- Data availability
- Author contributions
- Competing interests
- Disclaimer
- Special issue statement
- Acknowledgements
- Financial support
- Review statement
- References
- Abstract
- Introduction
- Methods
- Results
- Discussion
- Conclusions
- Appendix A: Selected characteristics of plant functional types and Pinus sylvestris parametrization
- Appendix B: Biomass, temperature, and maximum climatic water deficit maps for Berlin–Brandenburg
- Appendix C: Examples of SLA distributions
- Appendix D: Precipitation seasonality
- Code availability
- Data availability
- Author contributions
- Competing interests
- Disclaimer
- Special issue statement
- Acknowledgements
- Financial support
- Review statement
- References